Downloaded 14 times





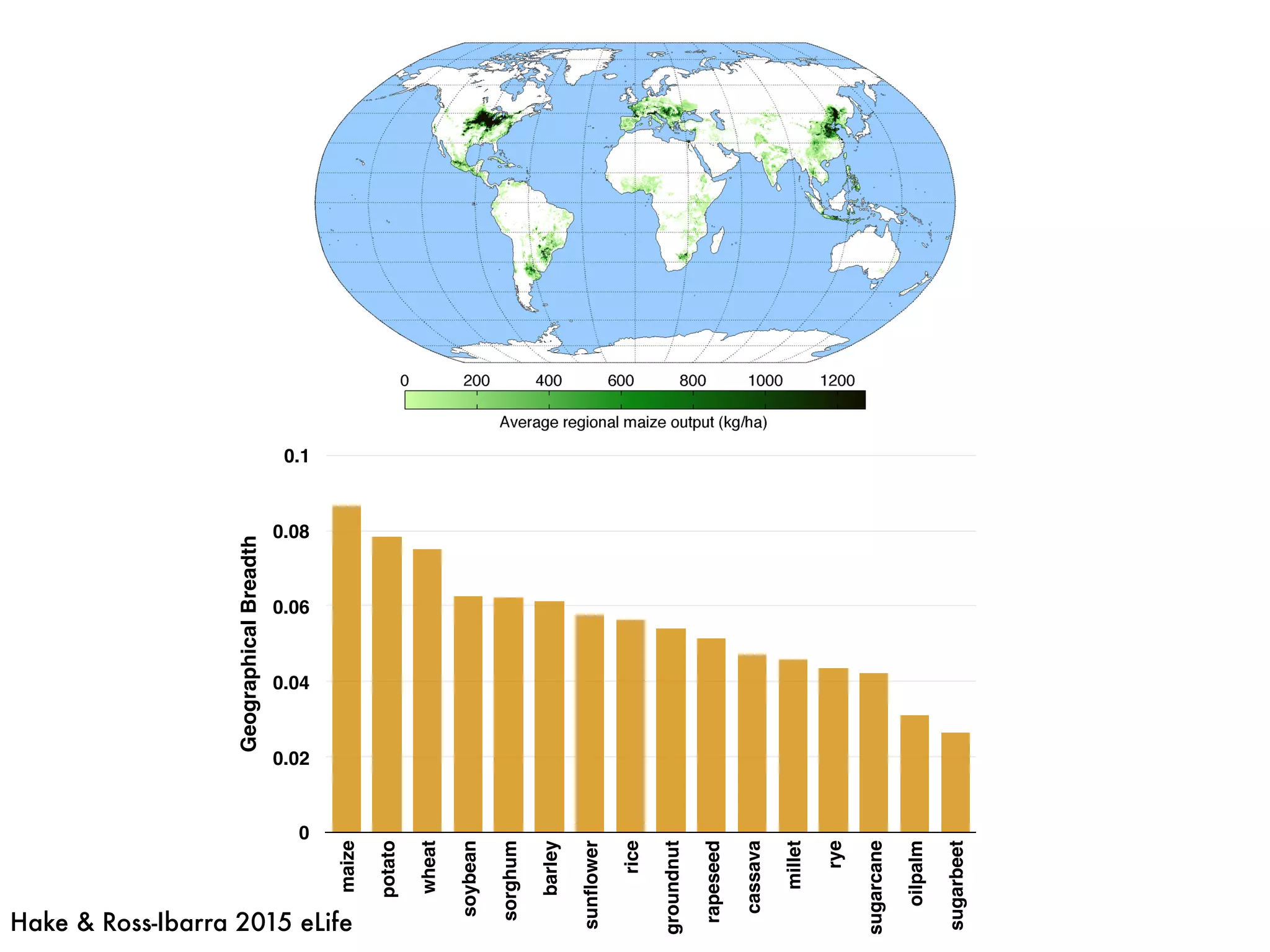
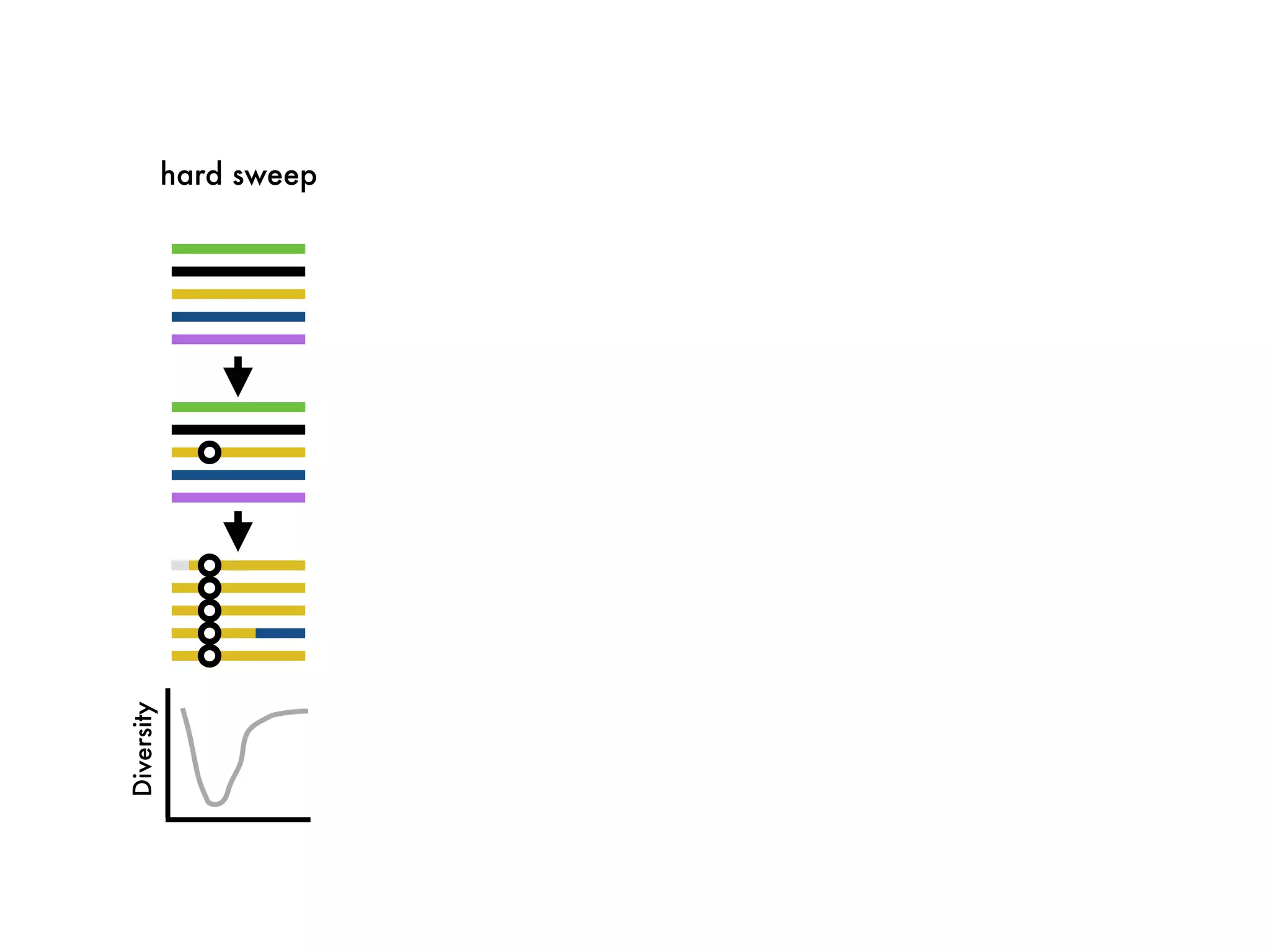
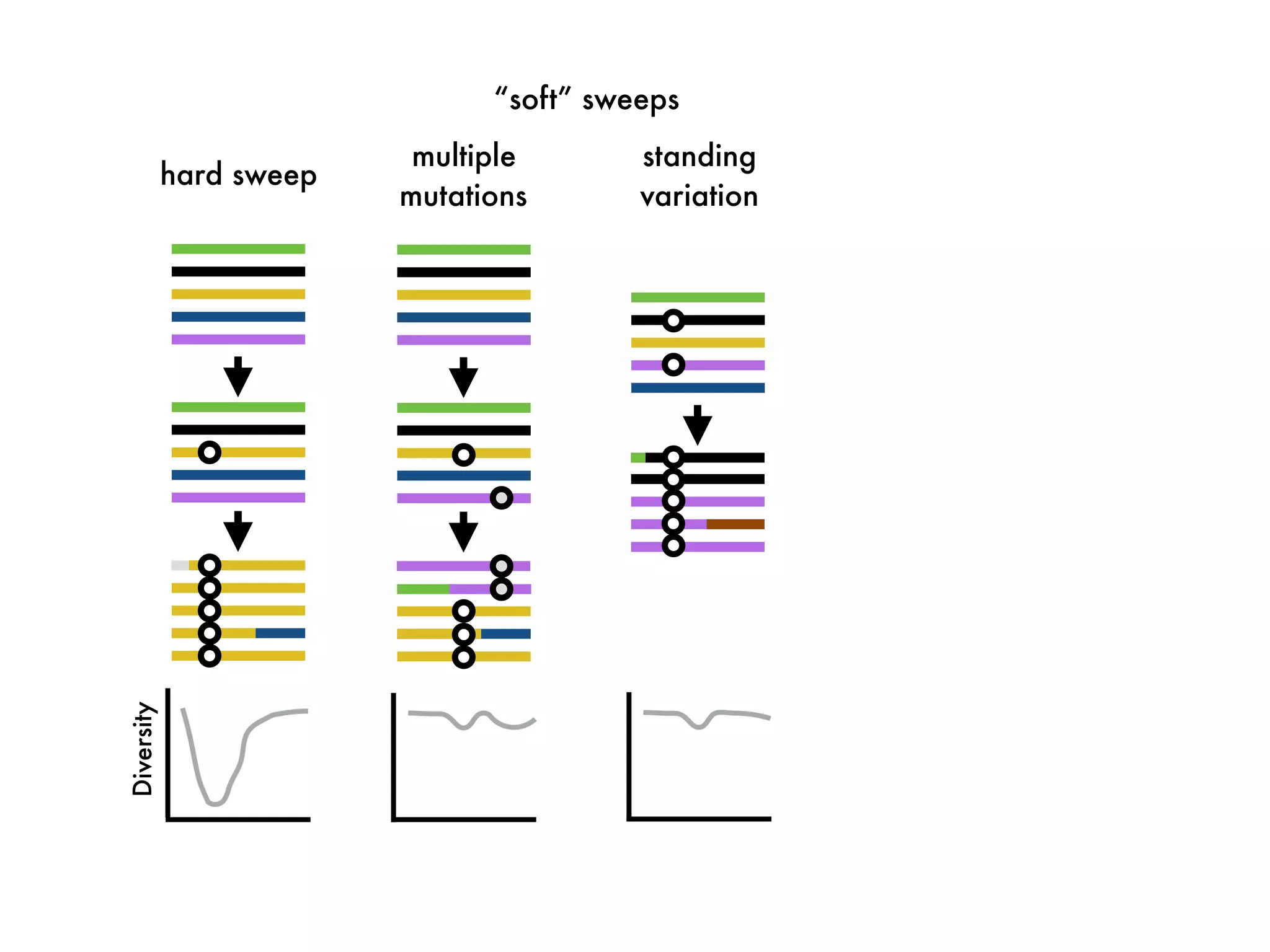
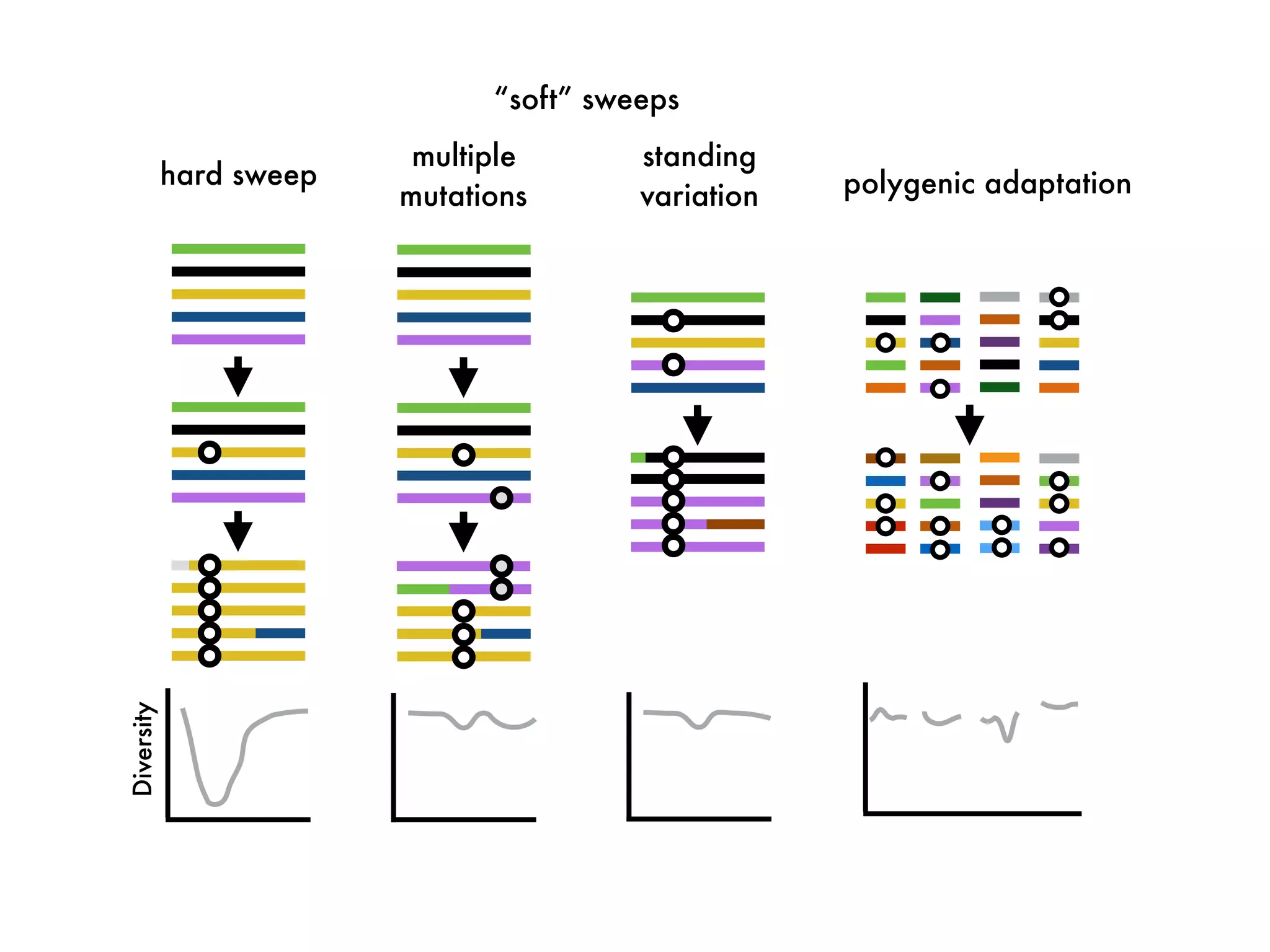
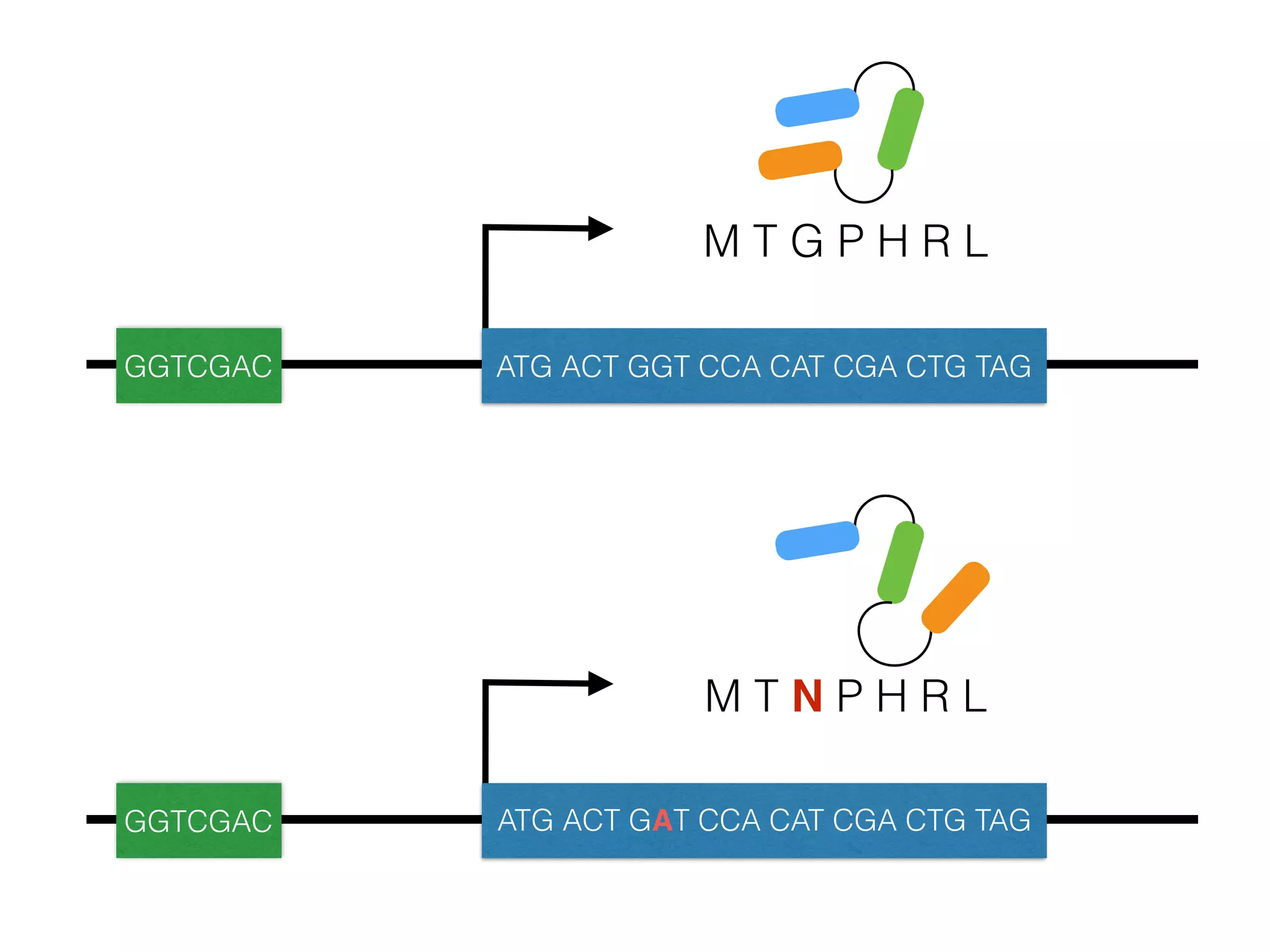
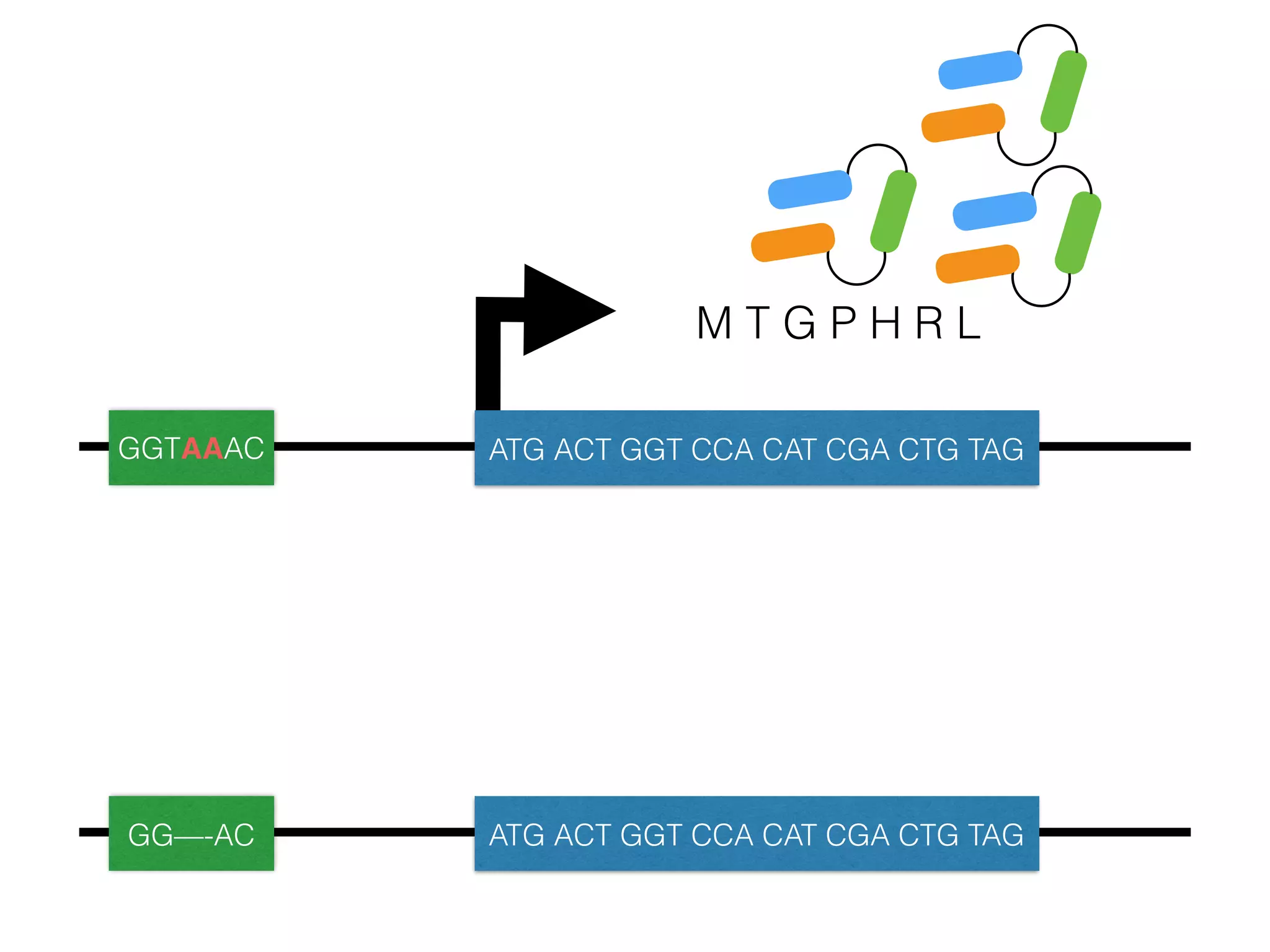
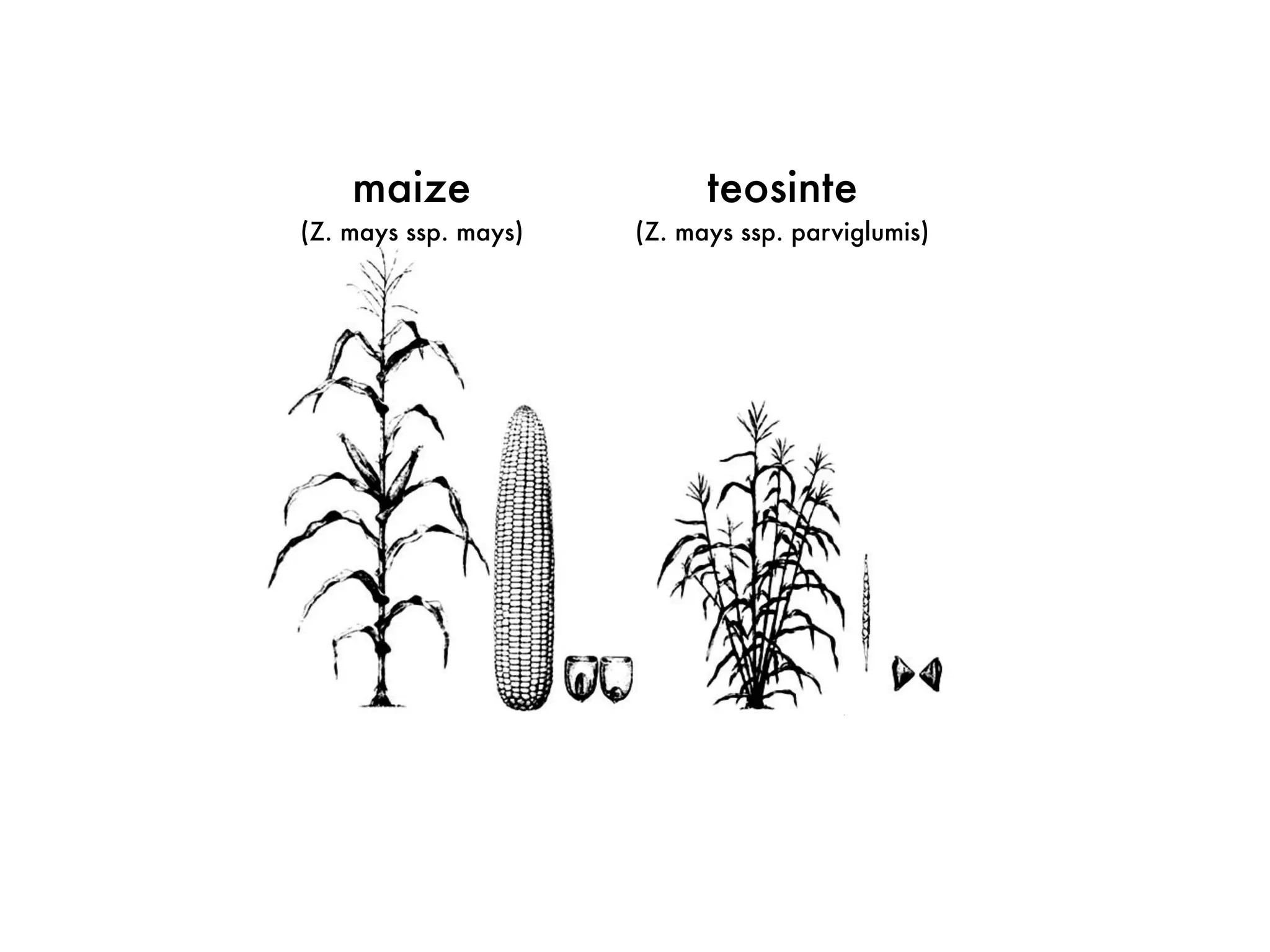
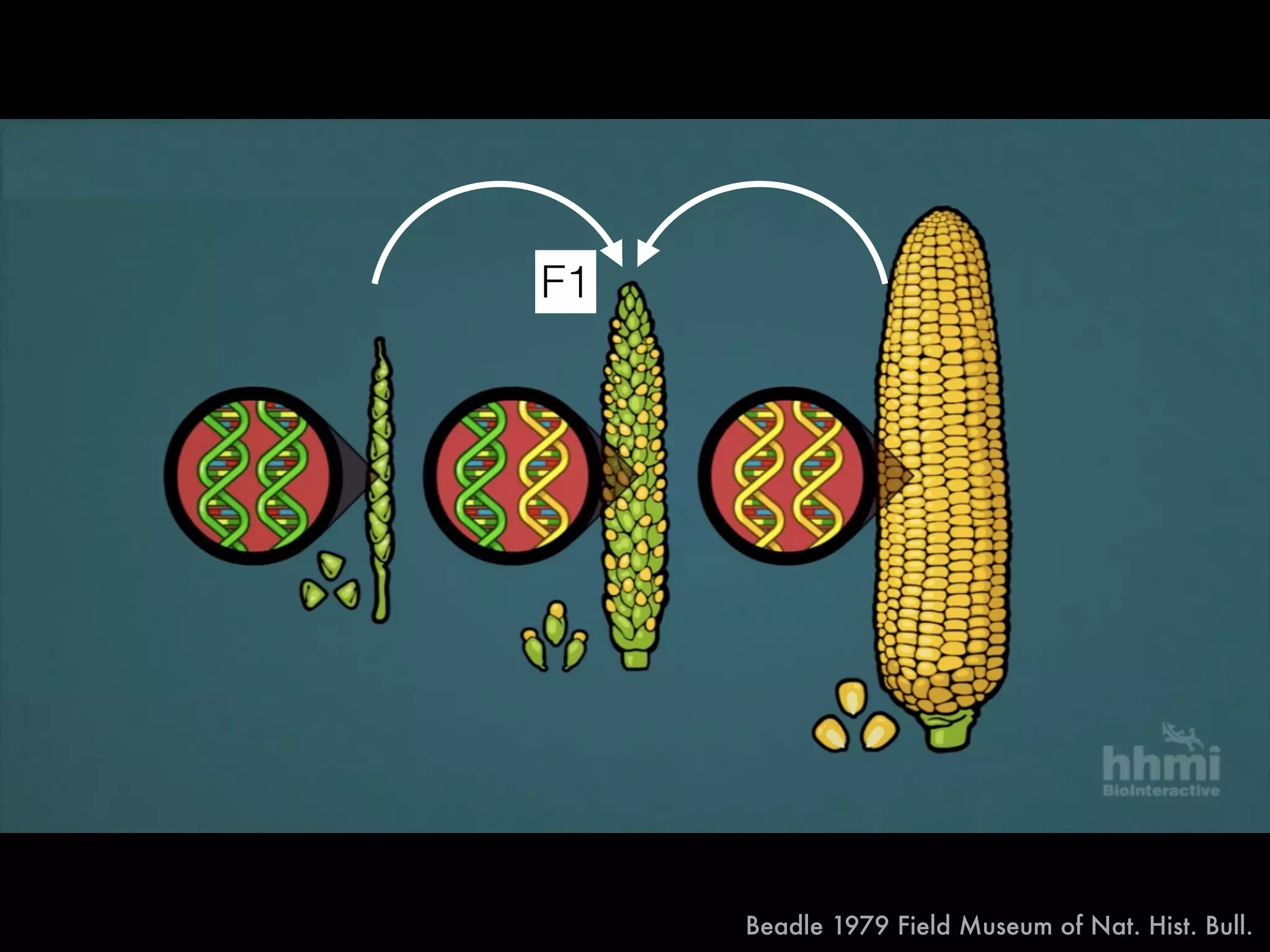
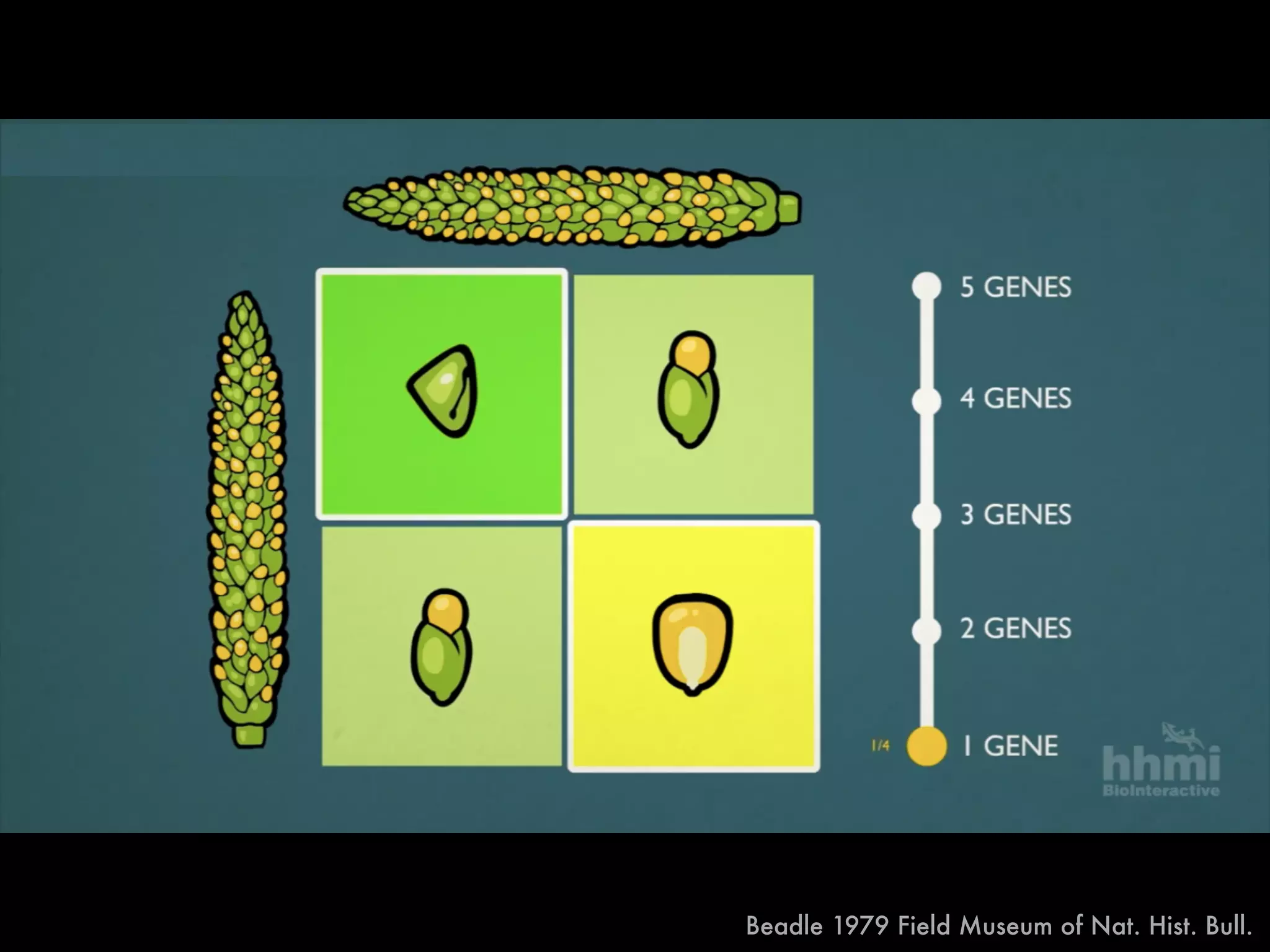
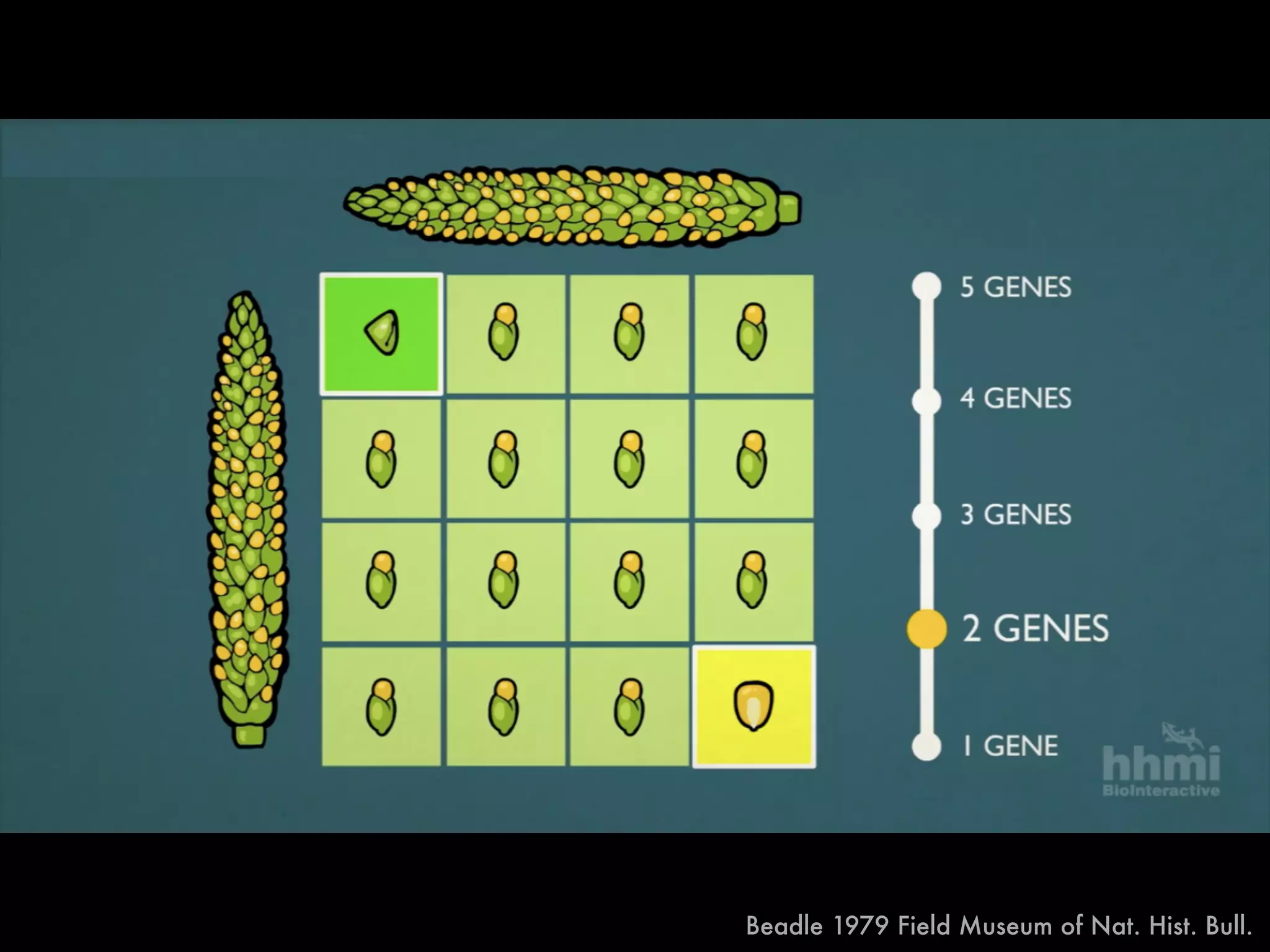

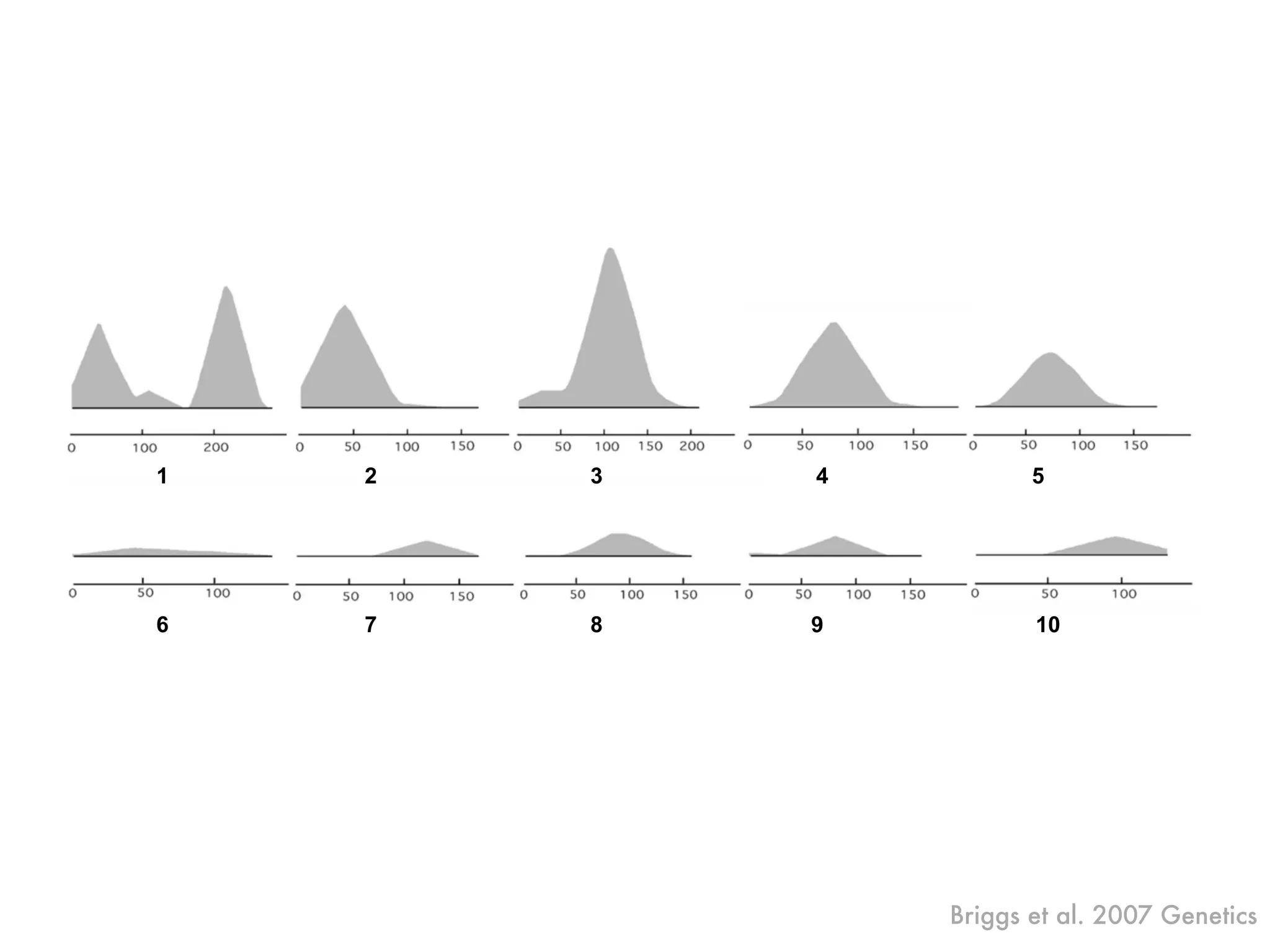
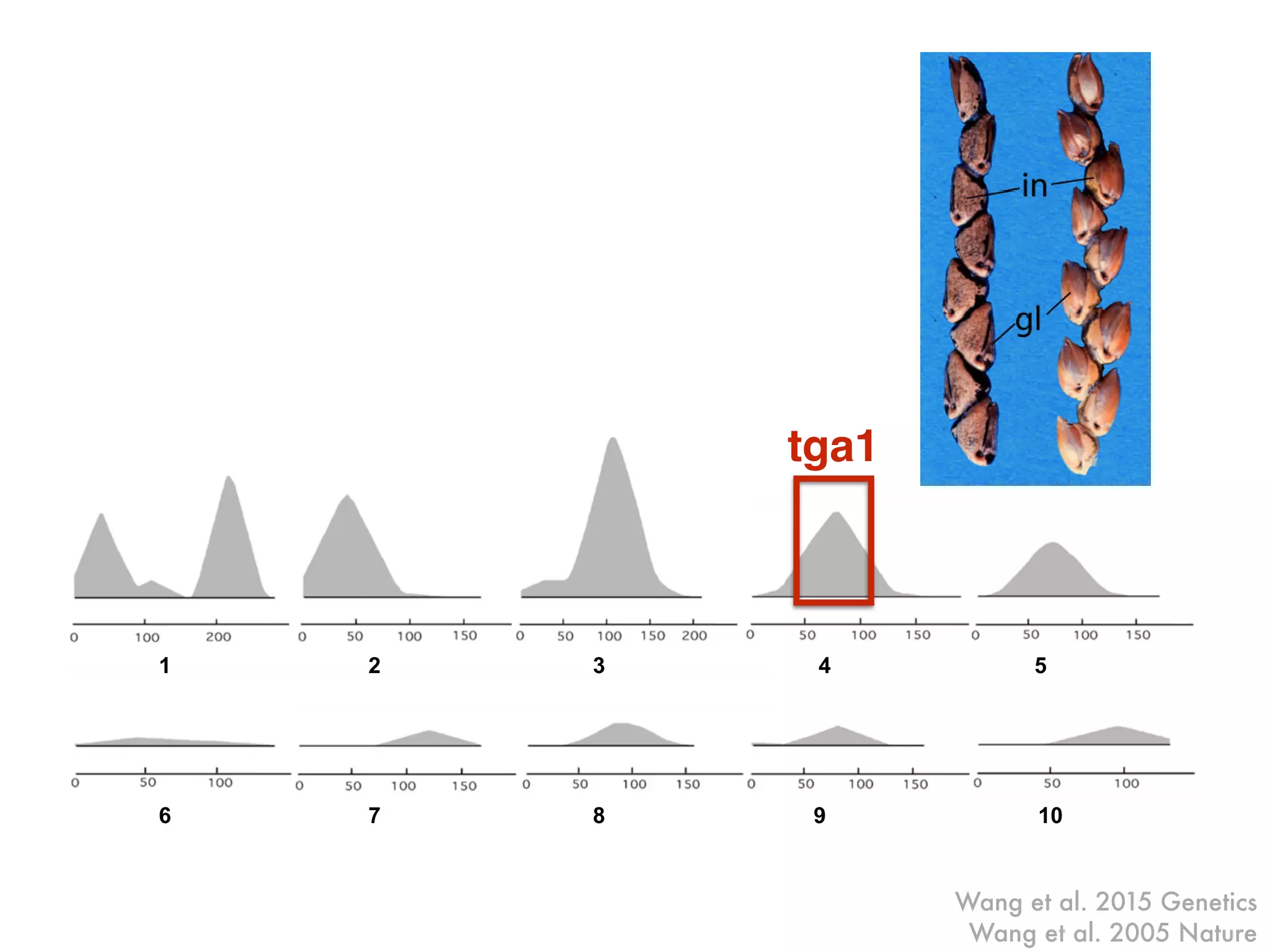
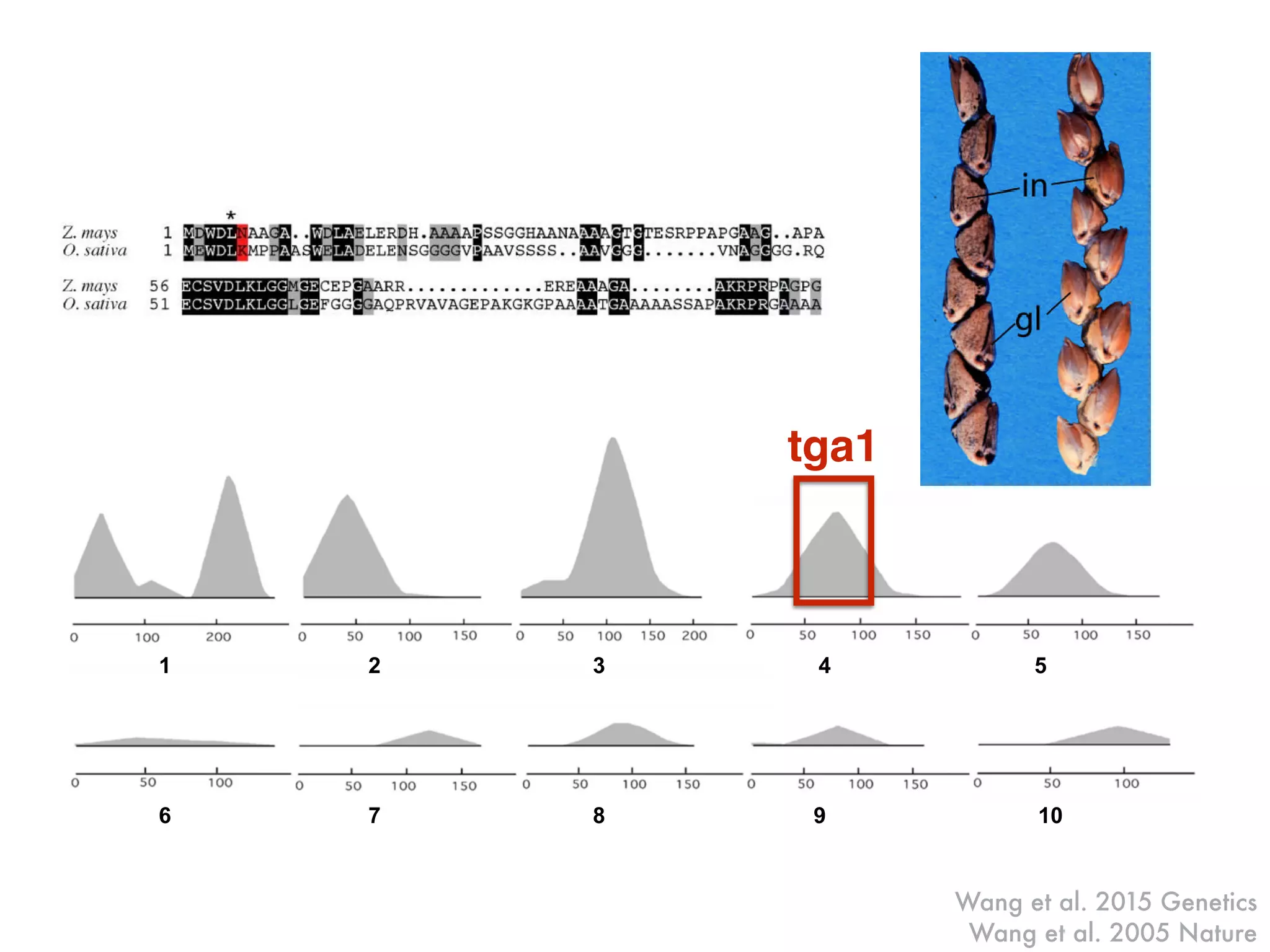
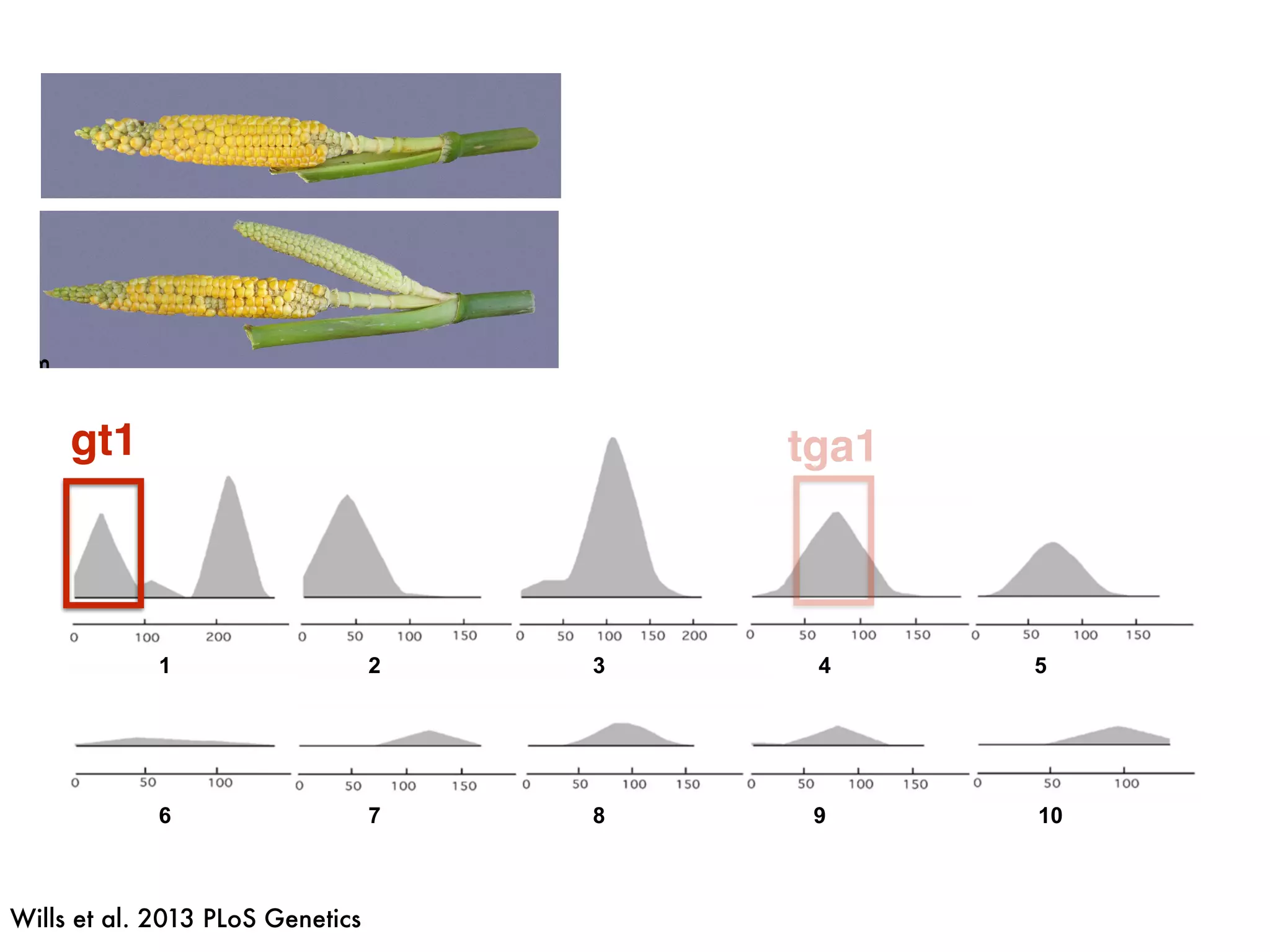
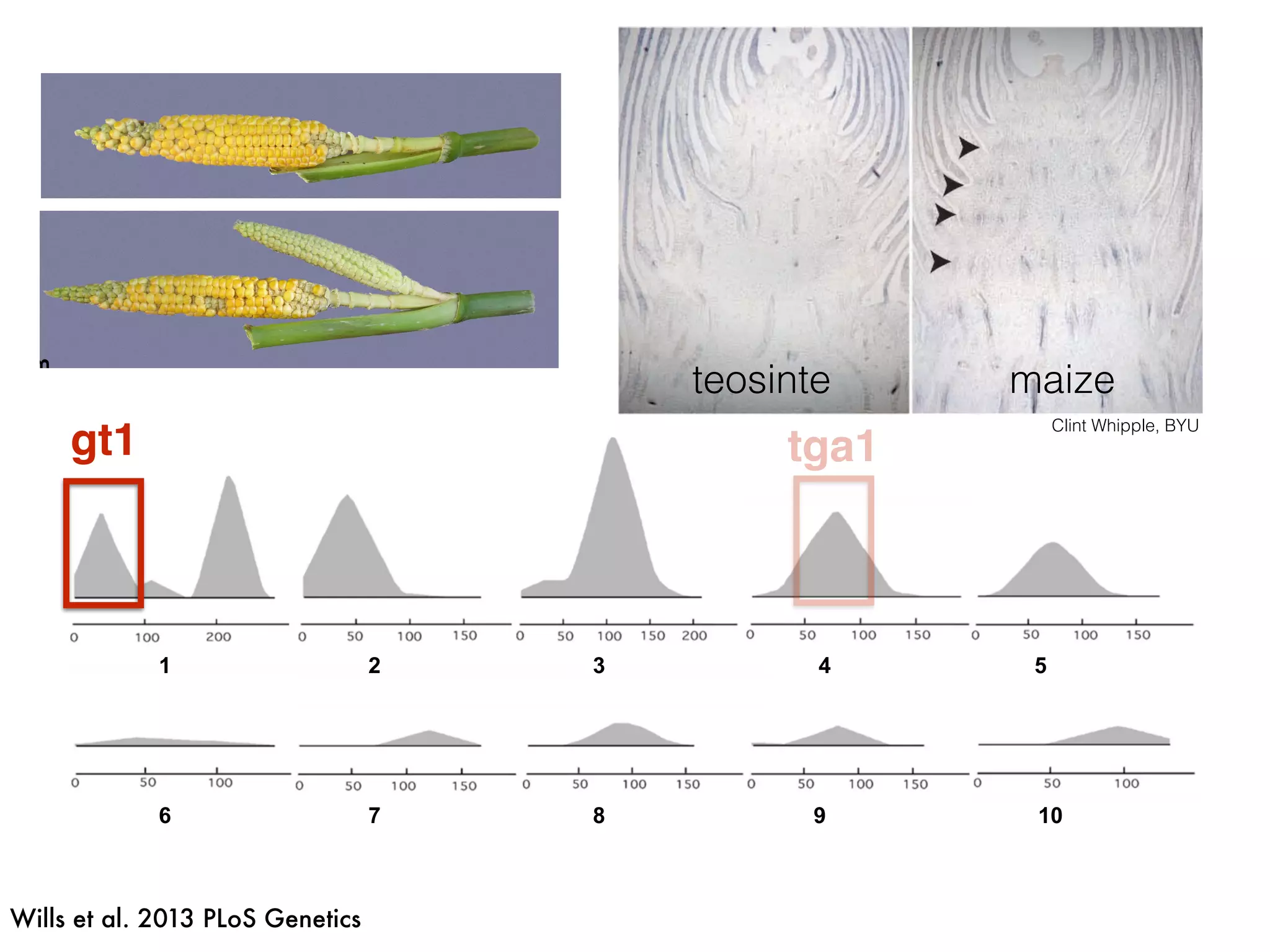
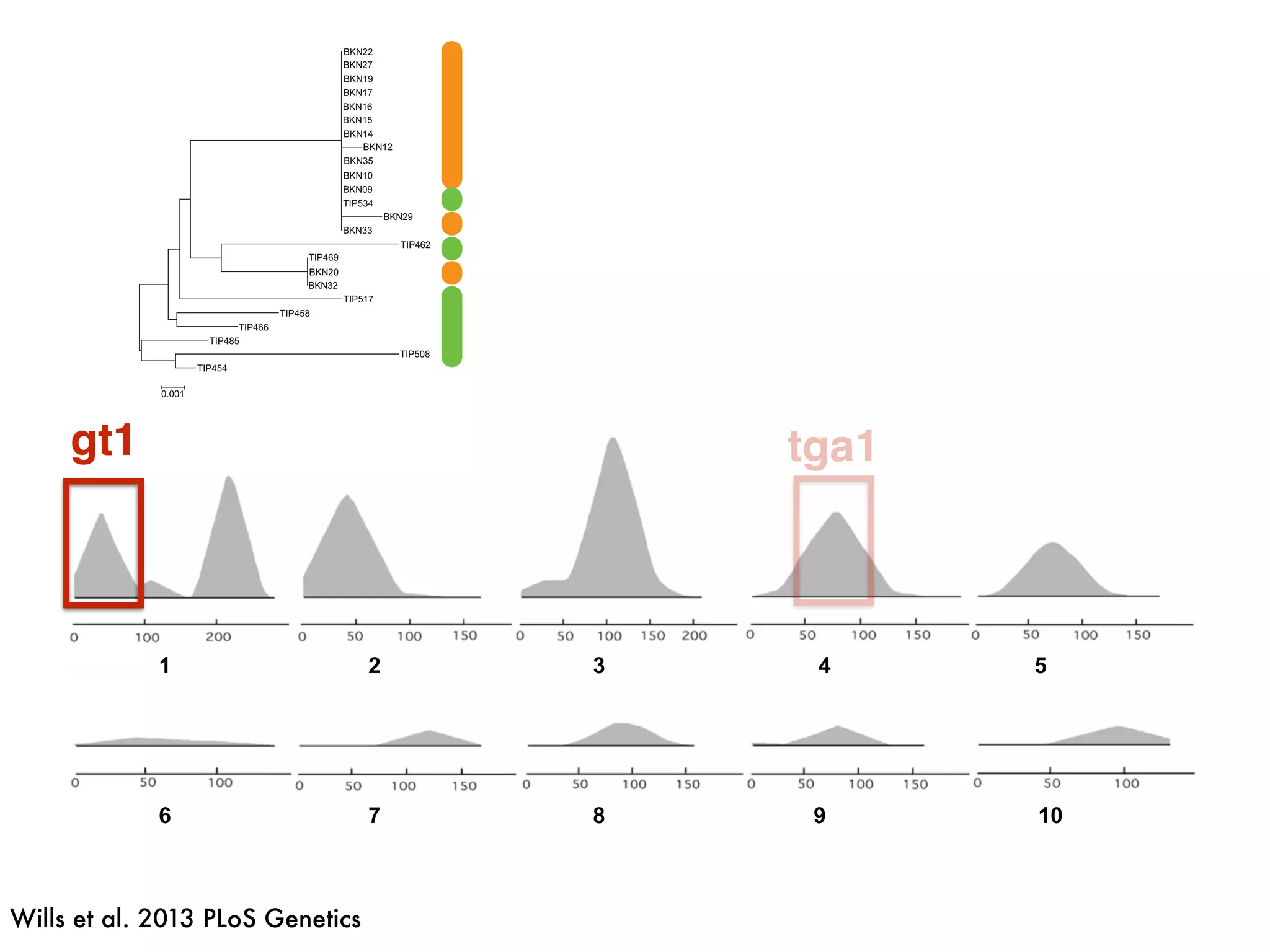
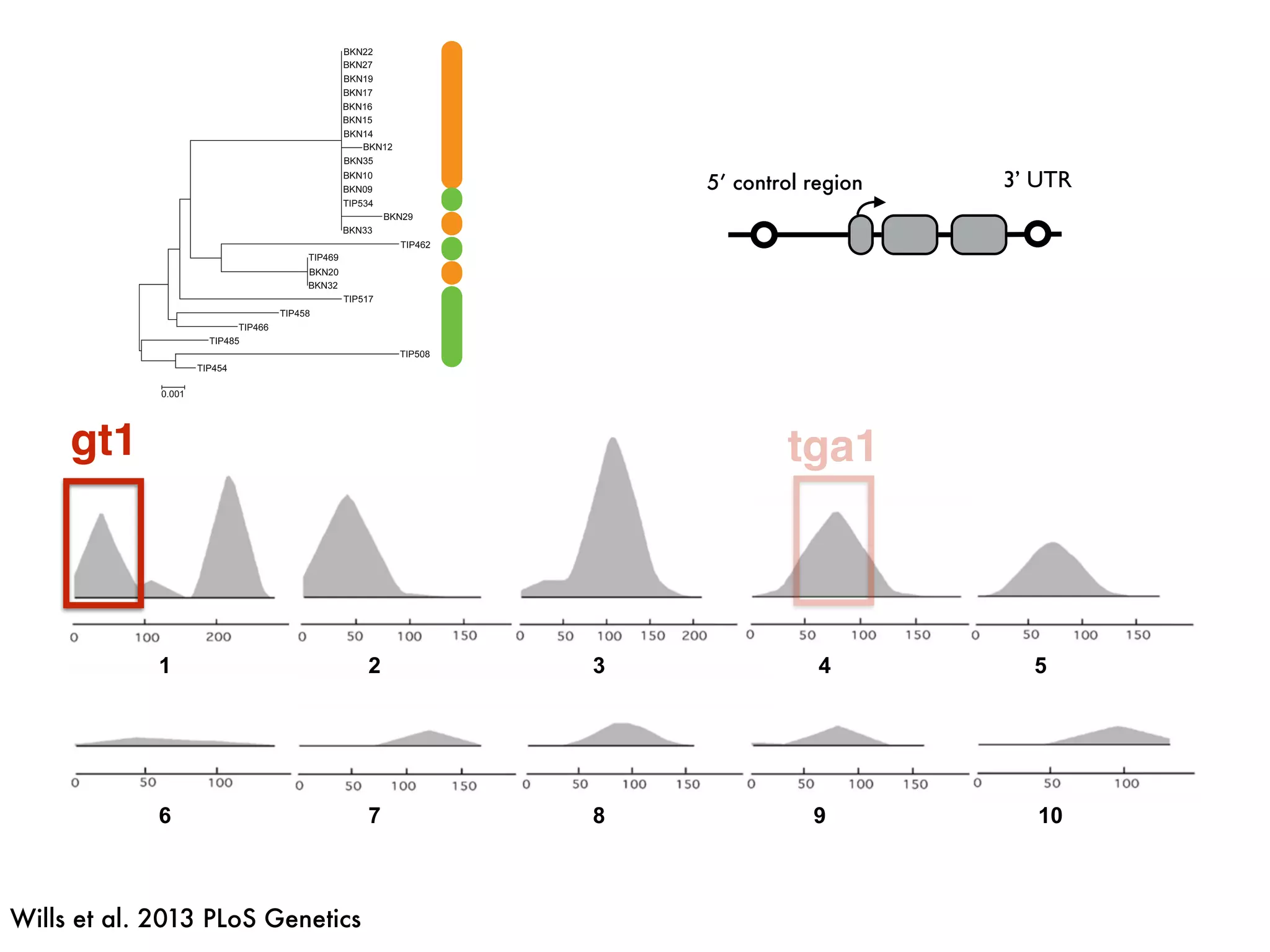
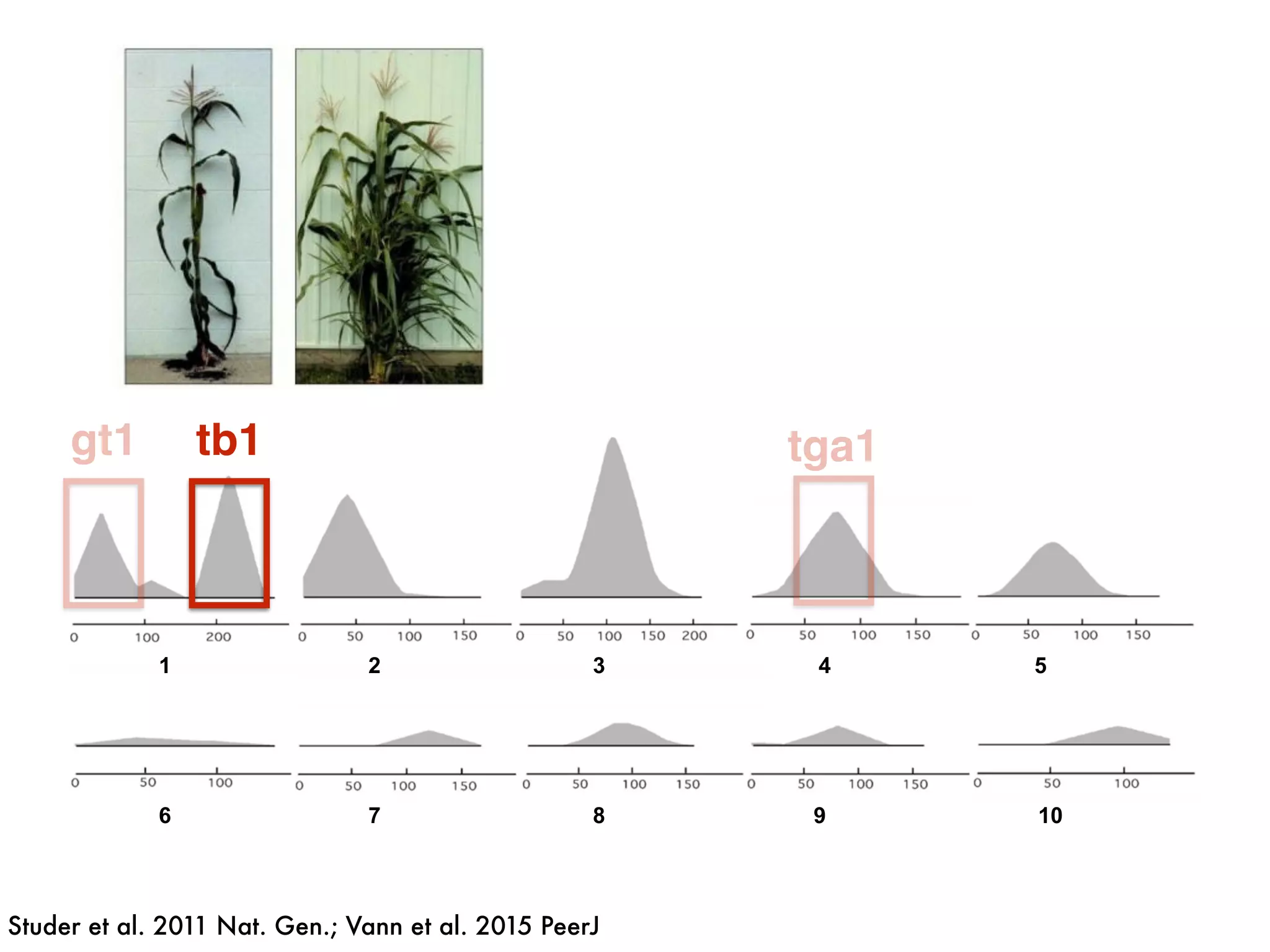

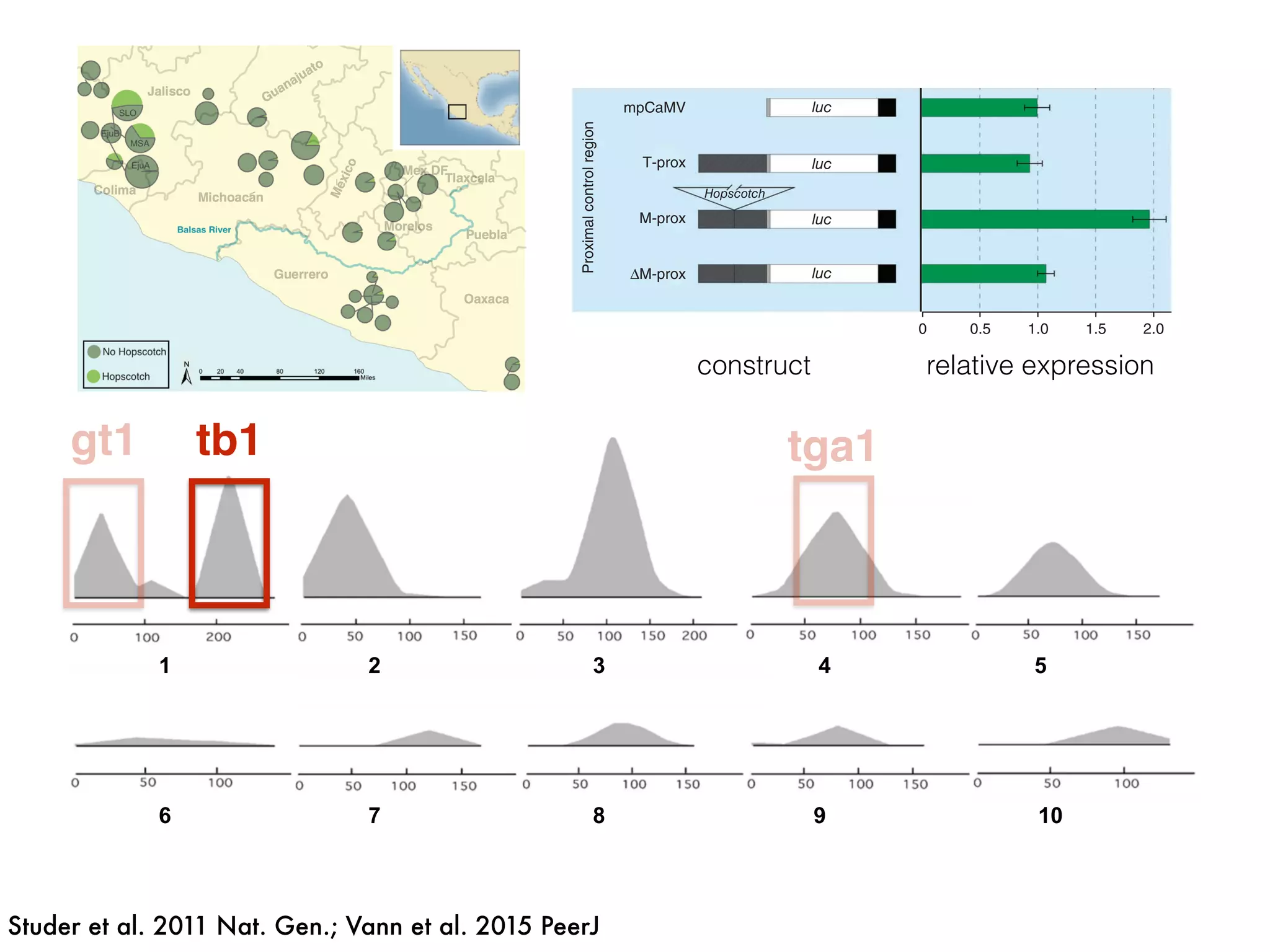
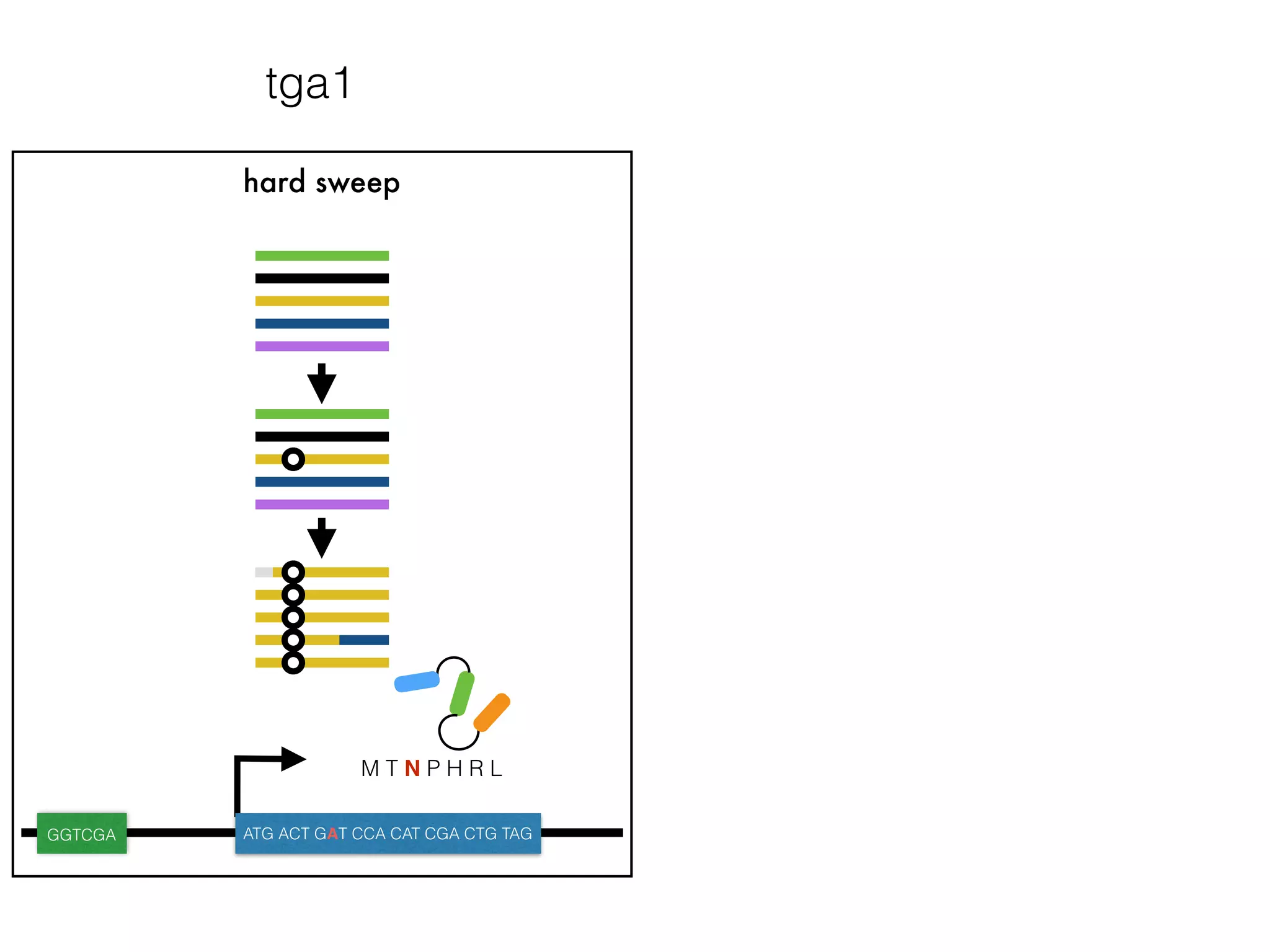
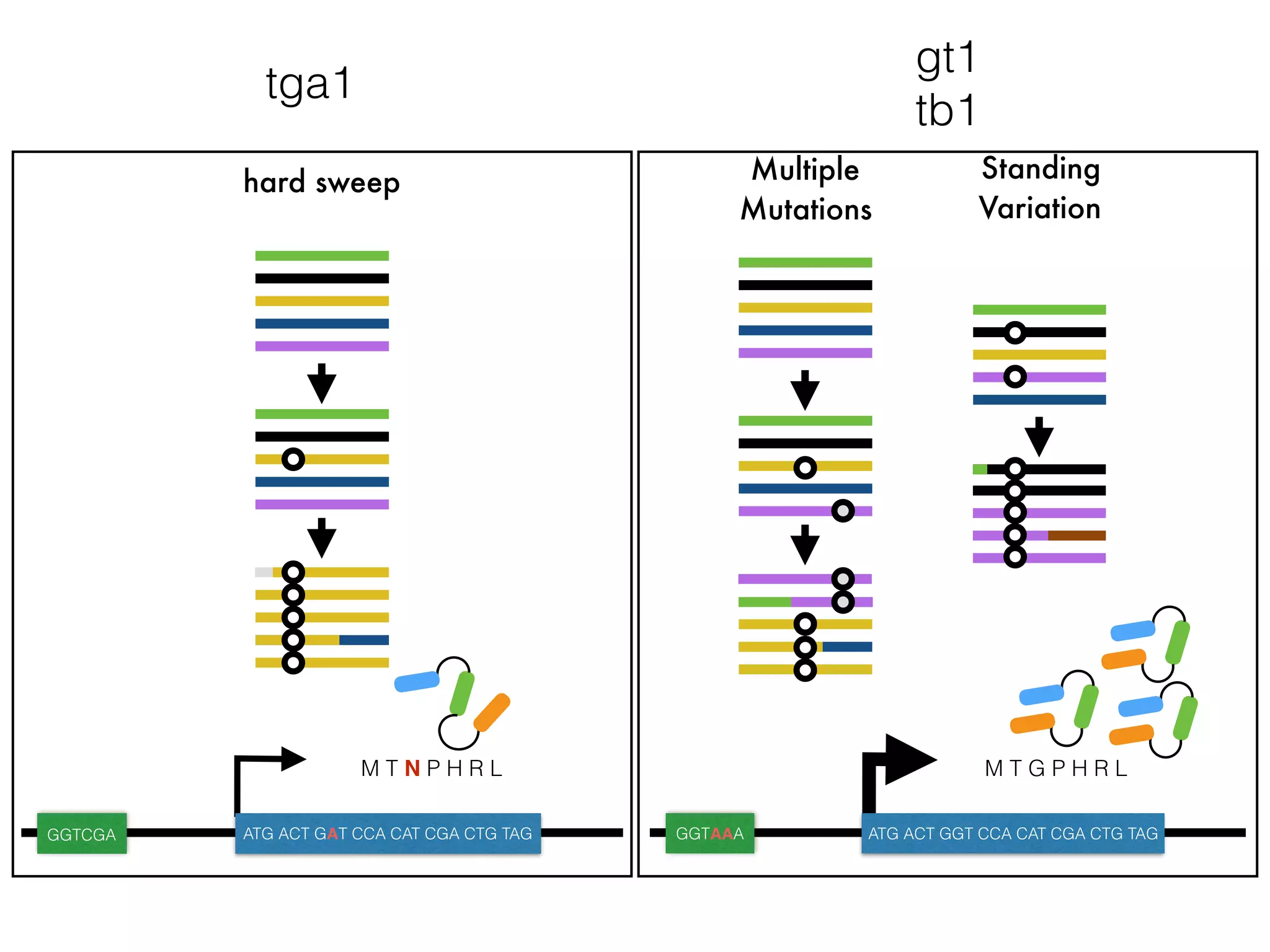
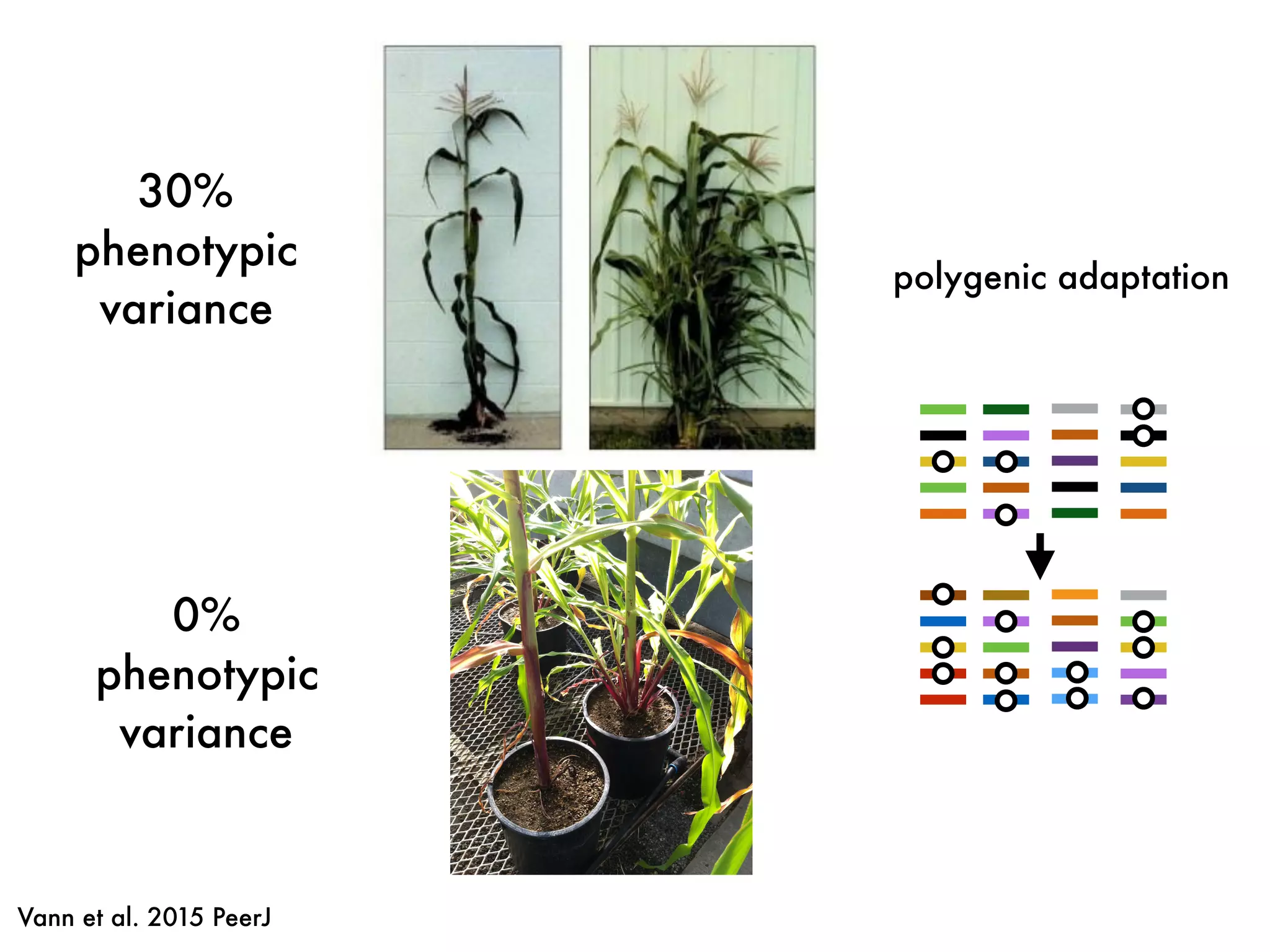
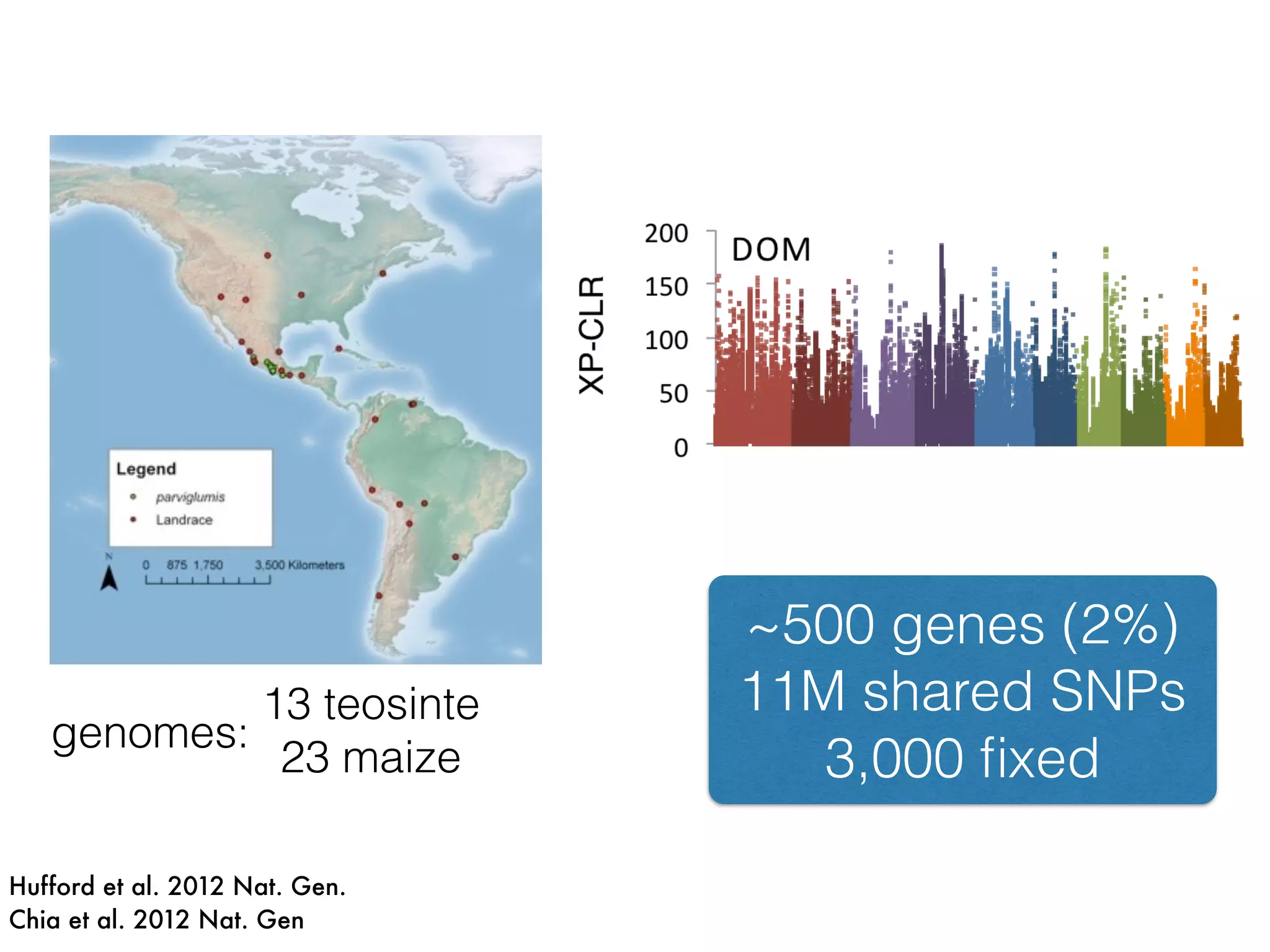
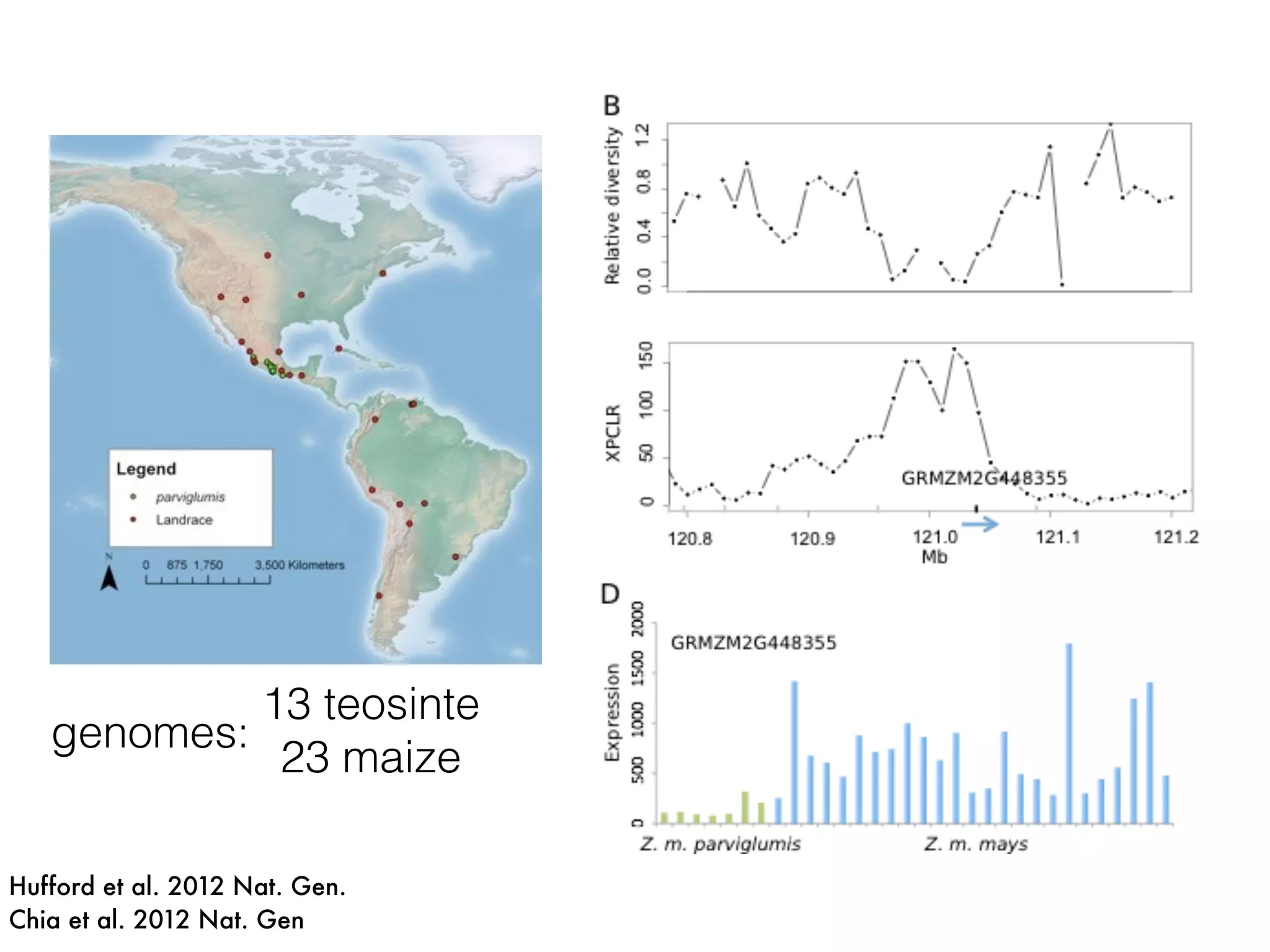
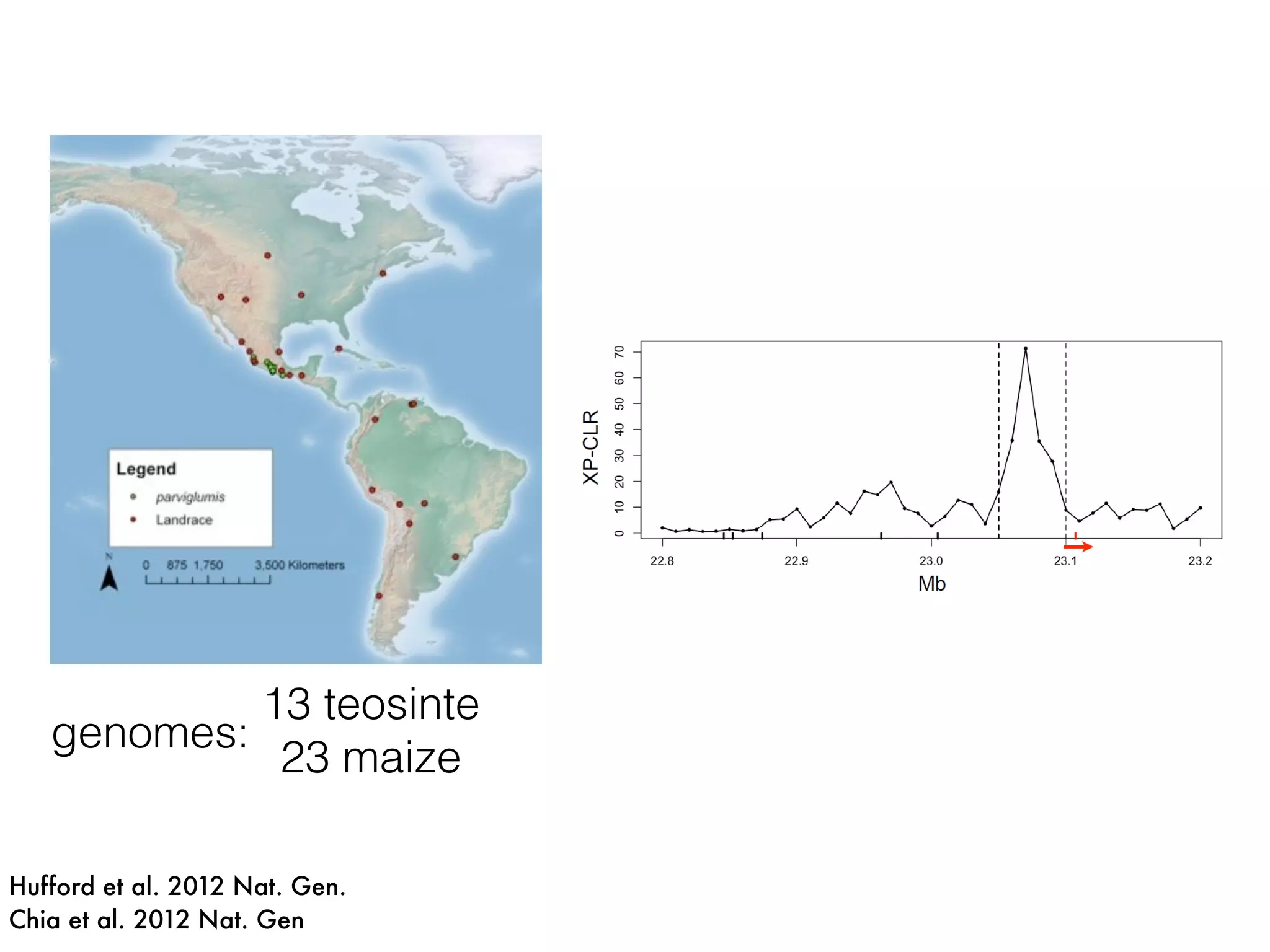
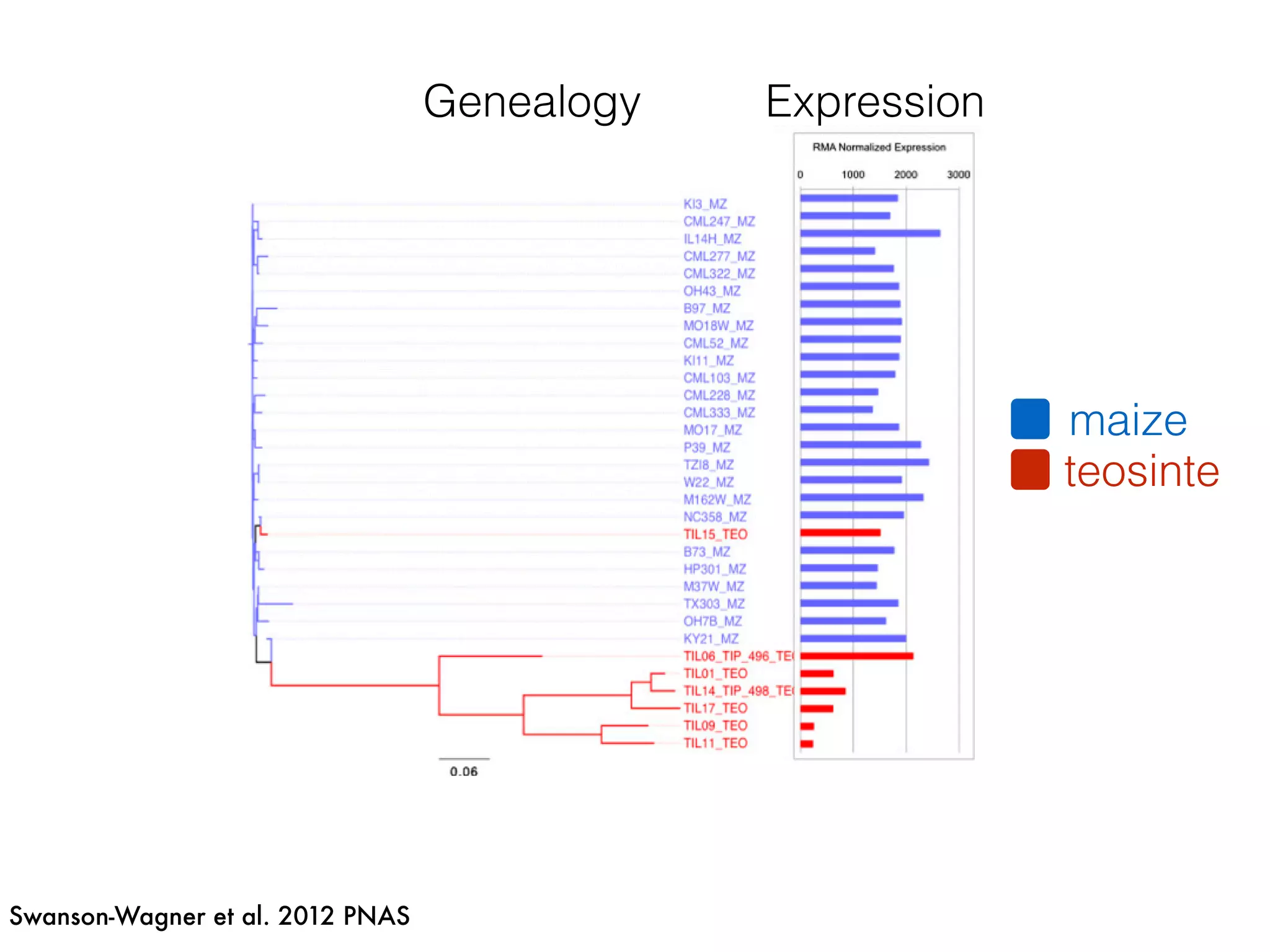
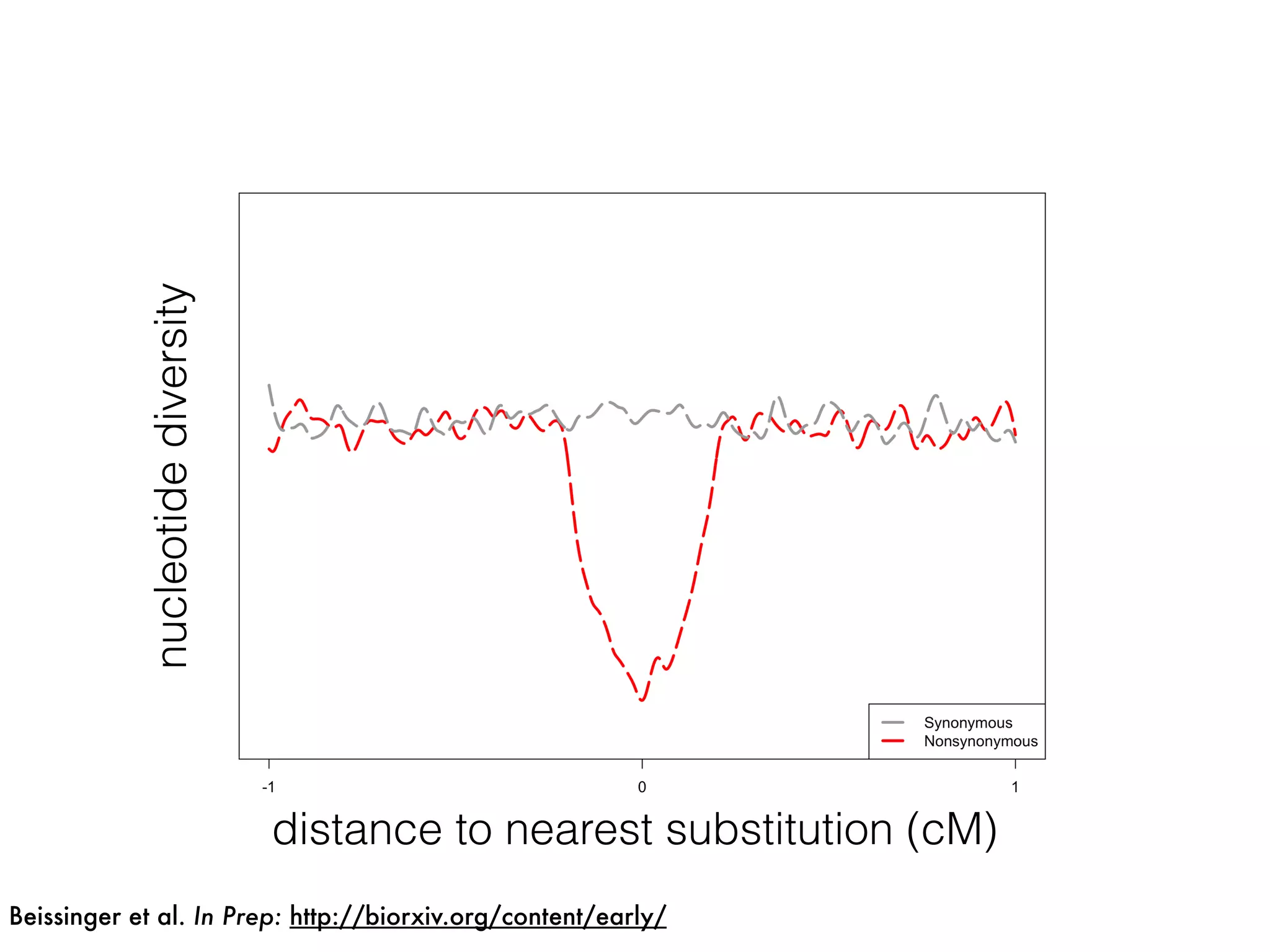
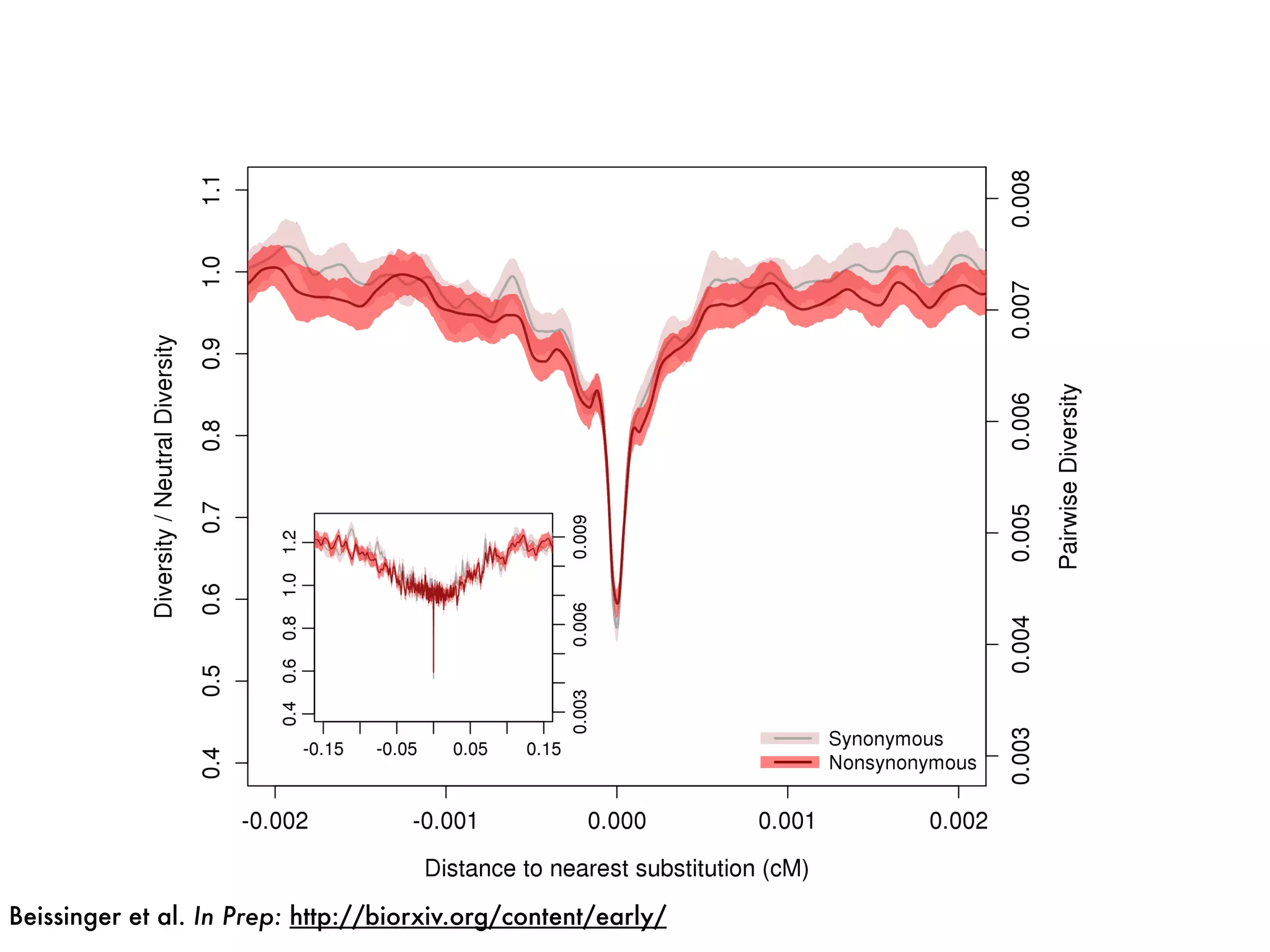
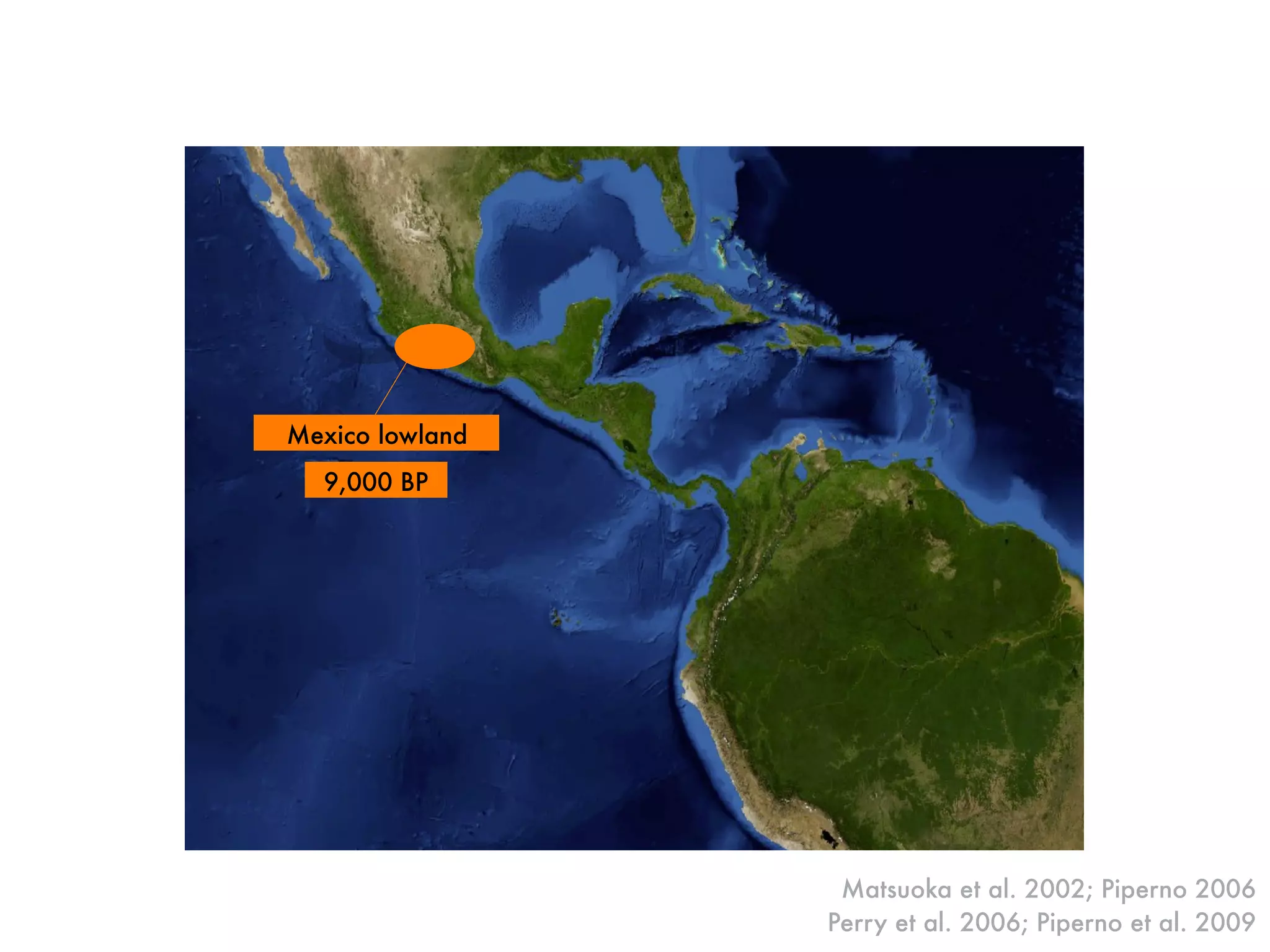
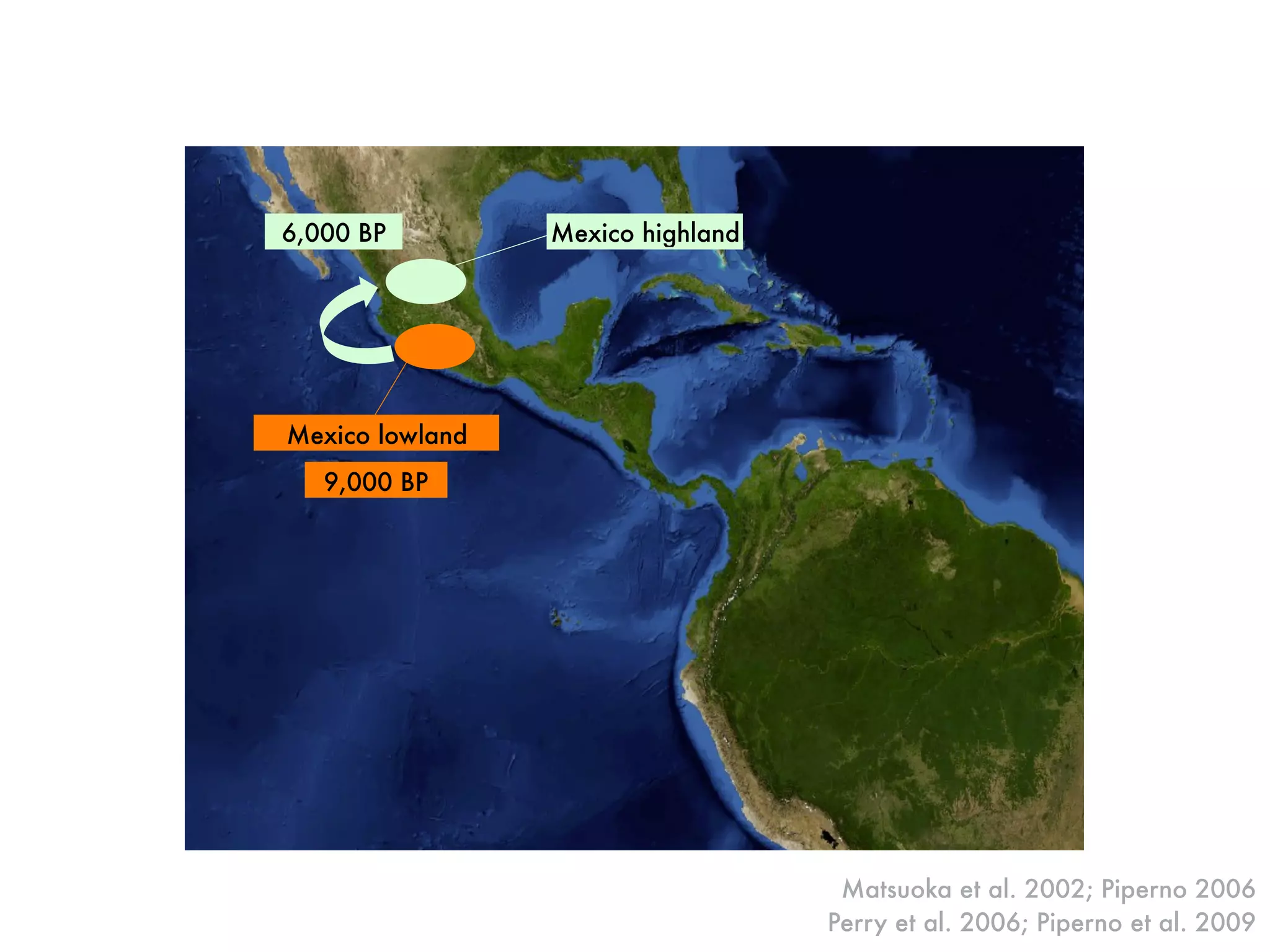
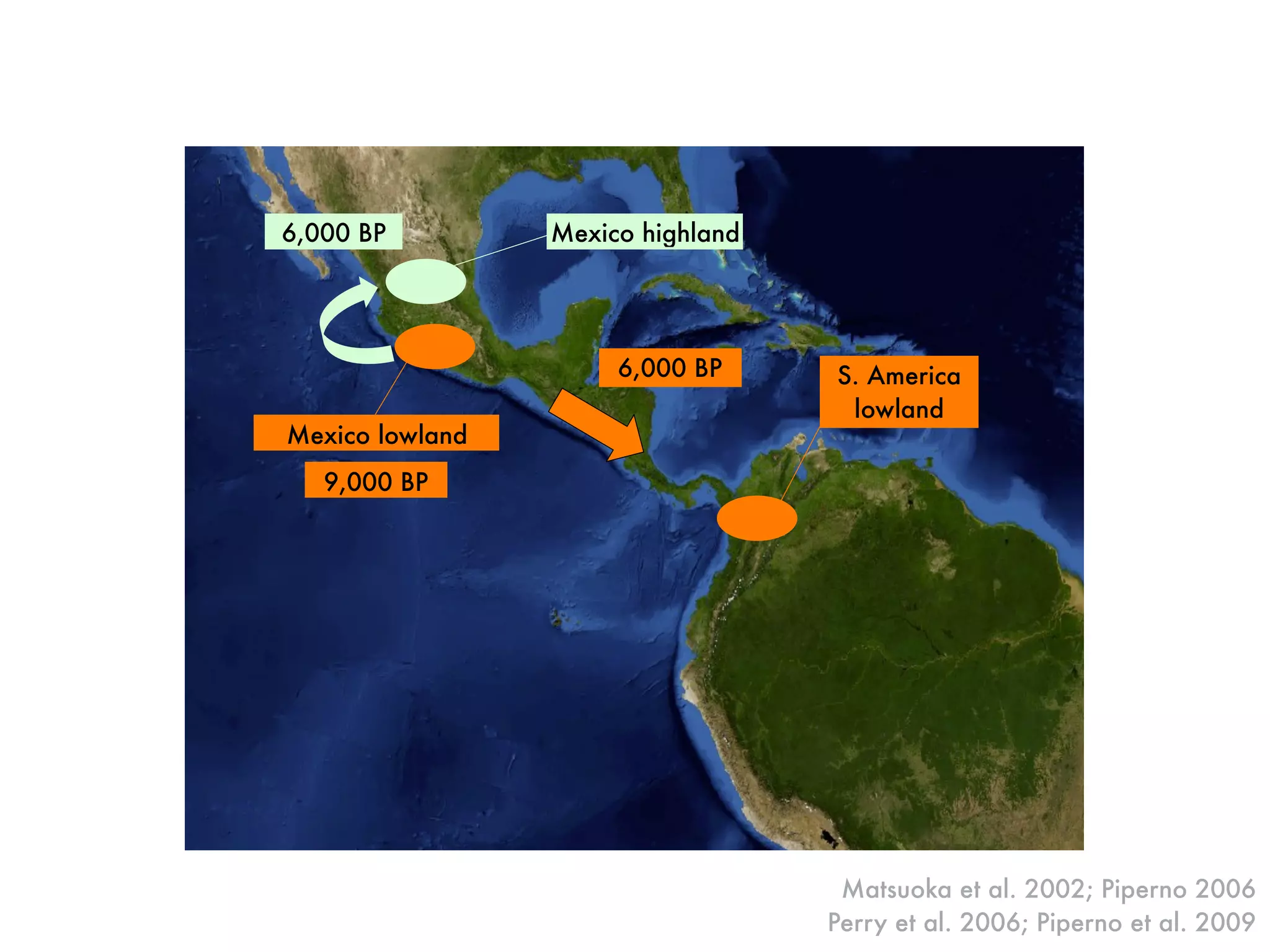
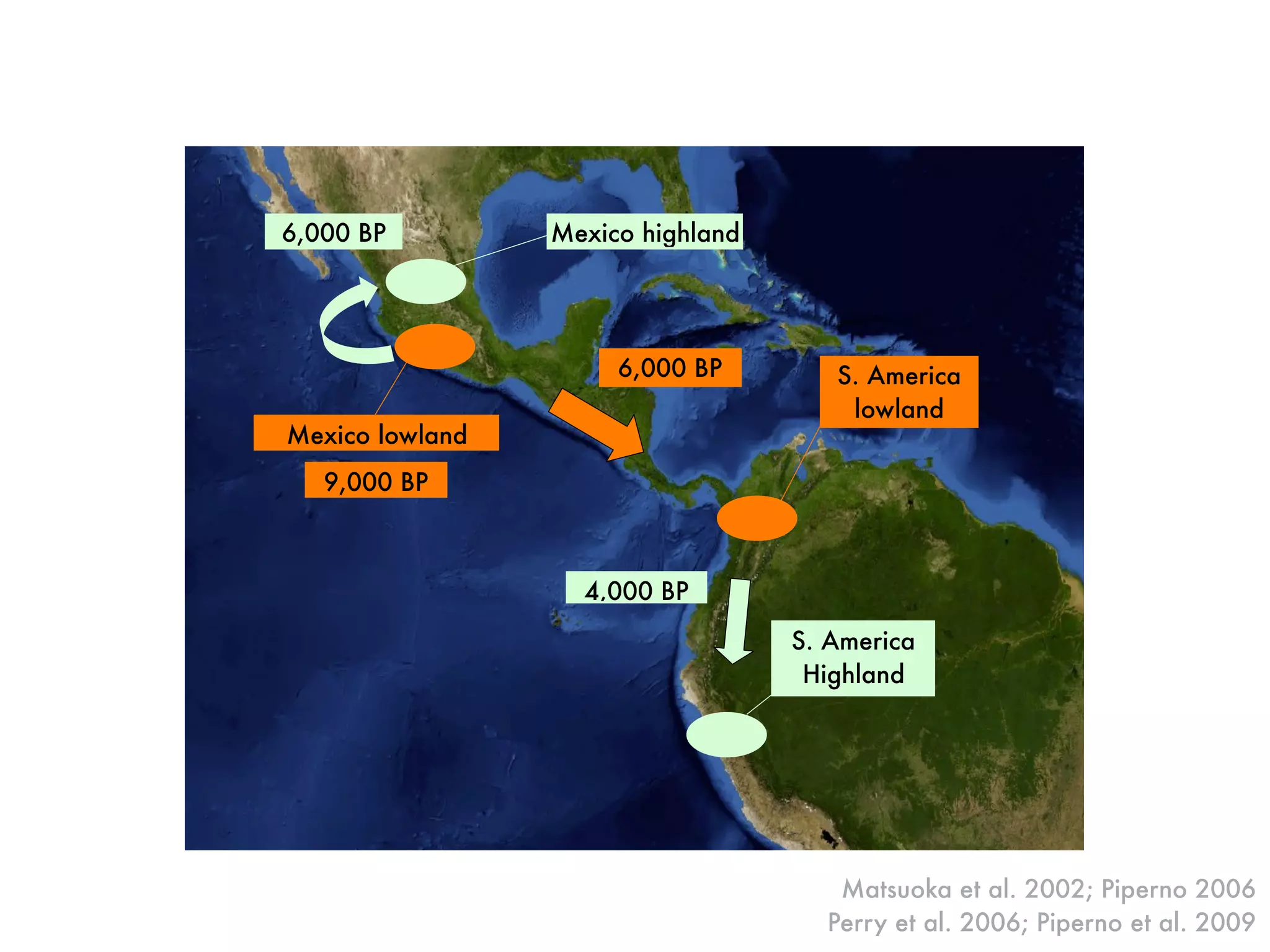

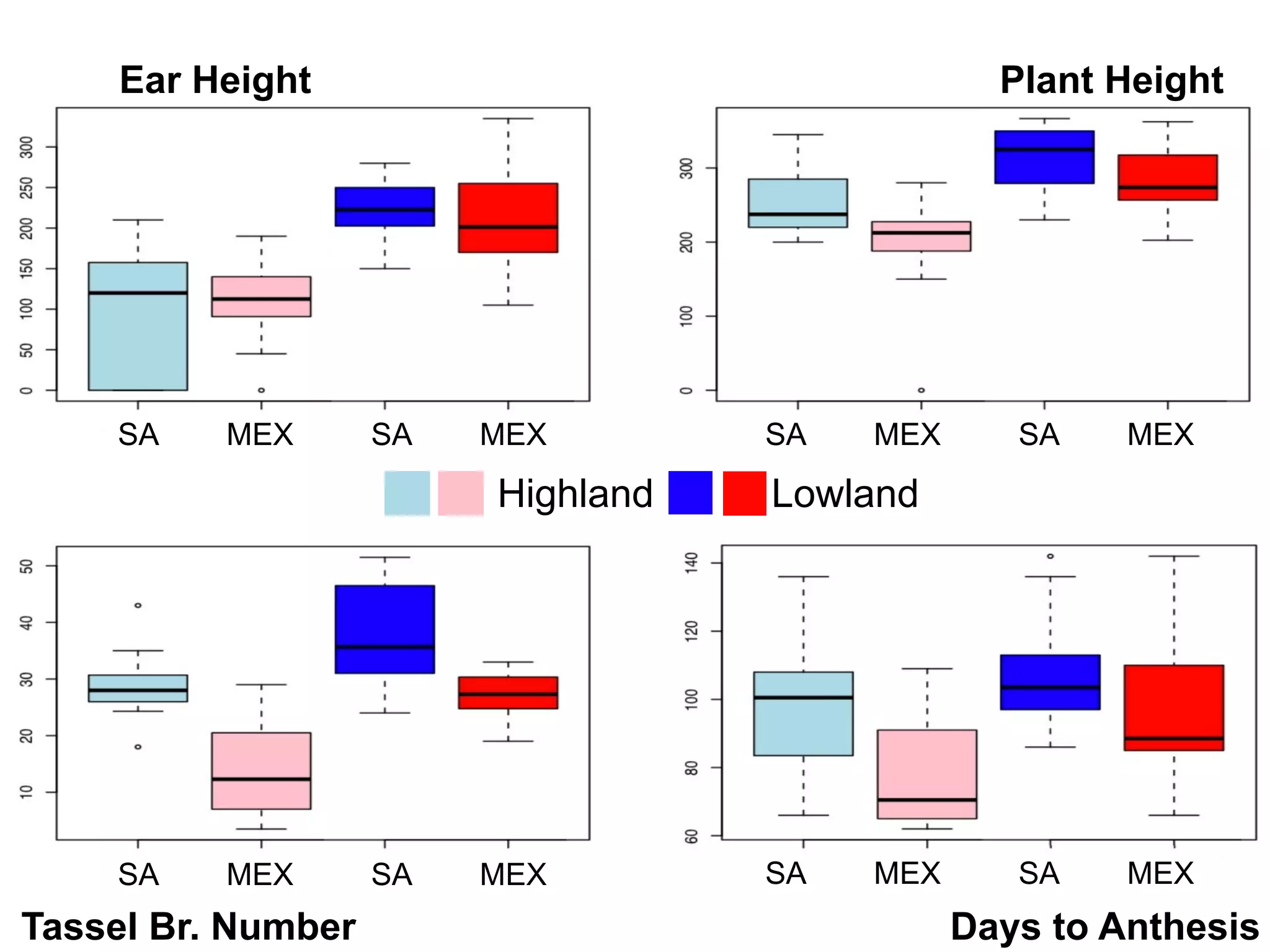
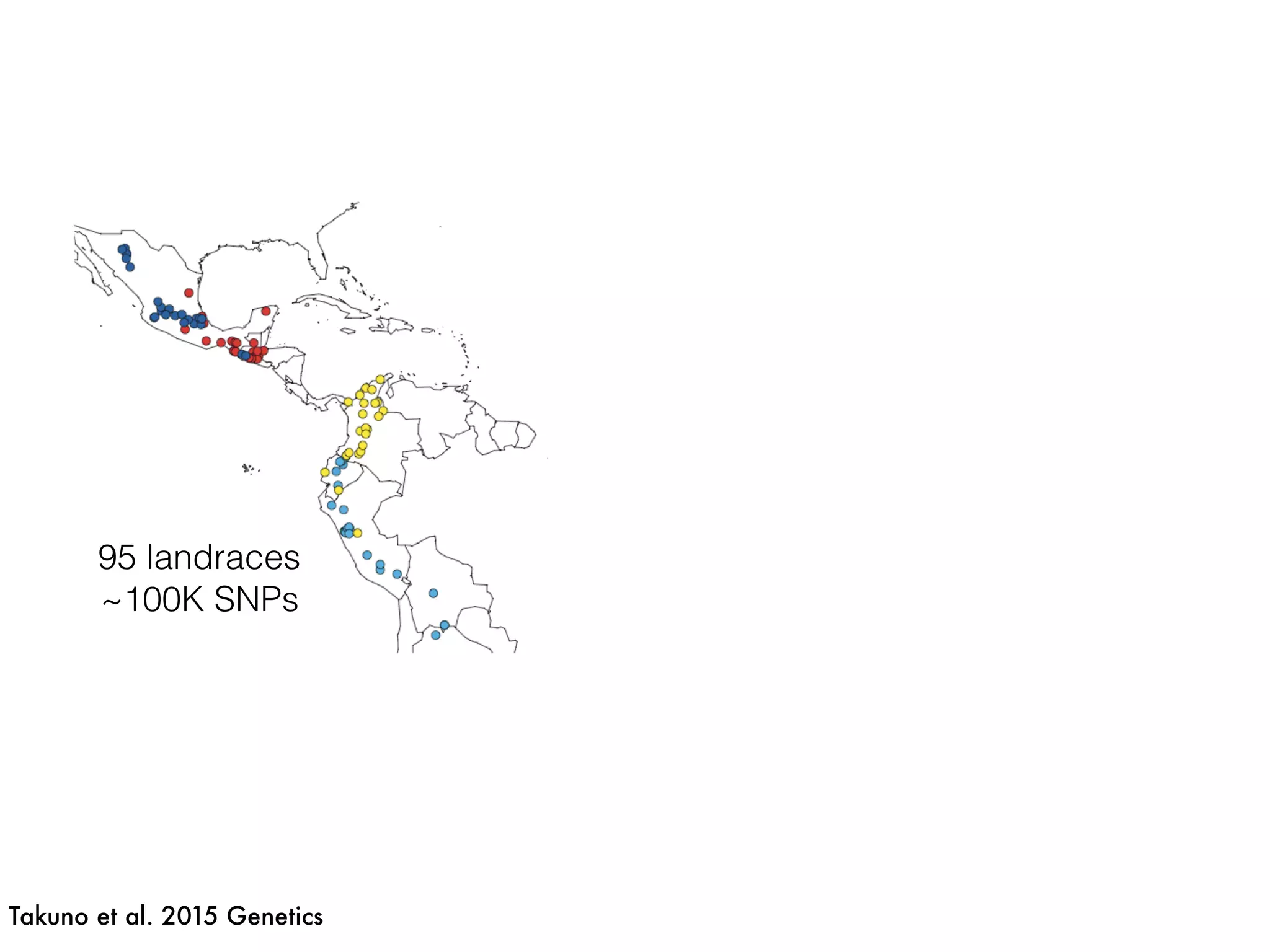
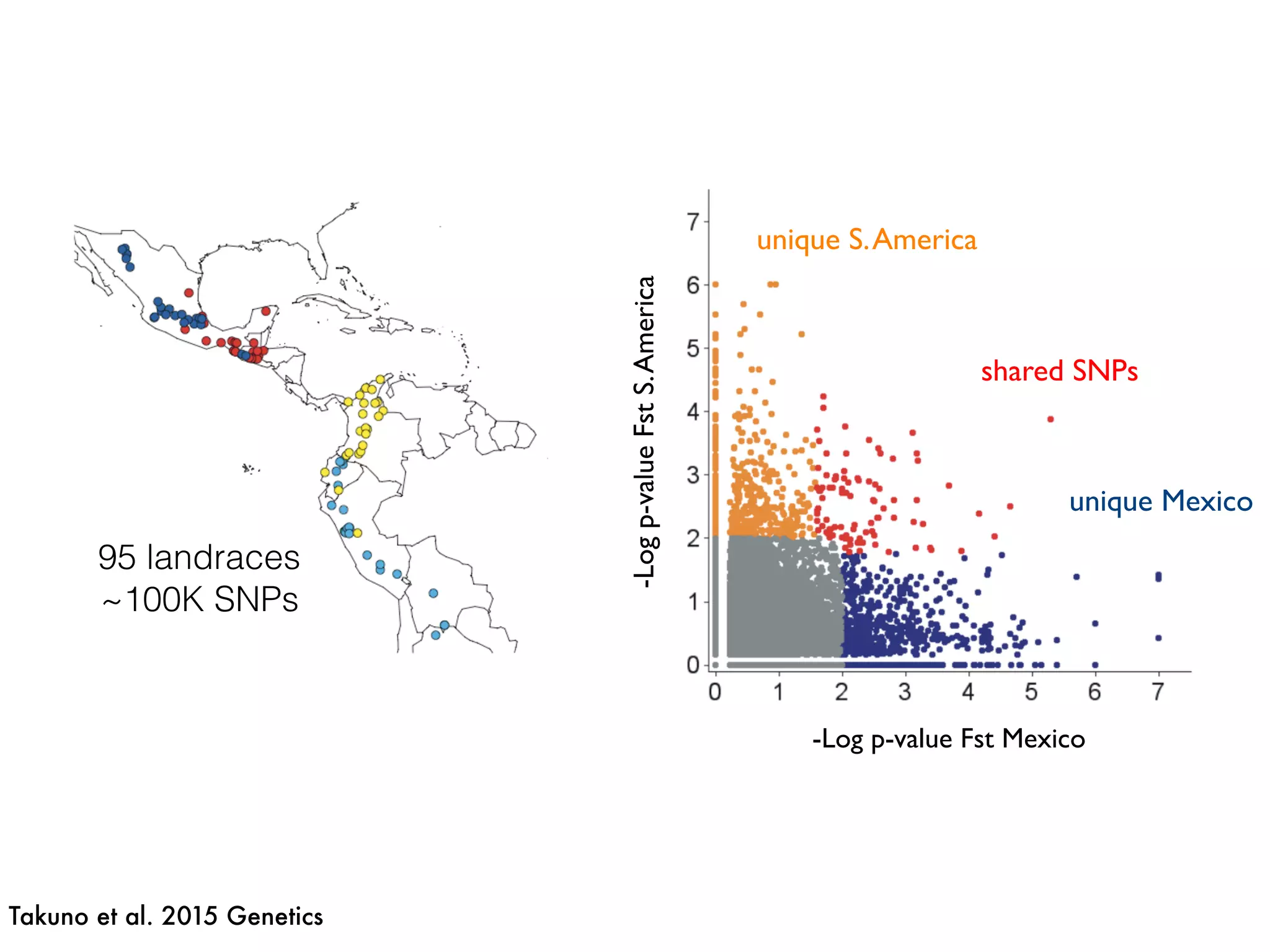
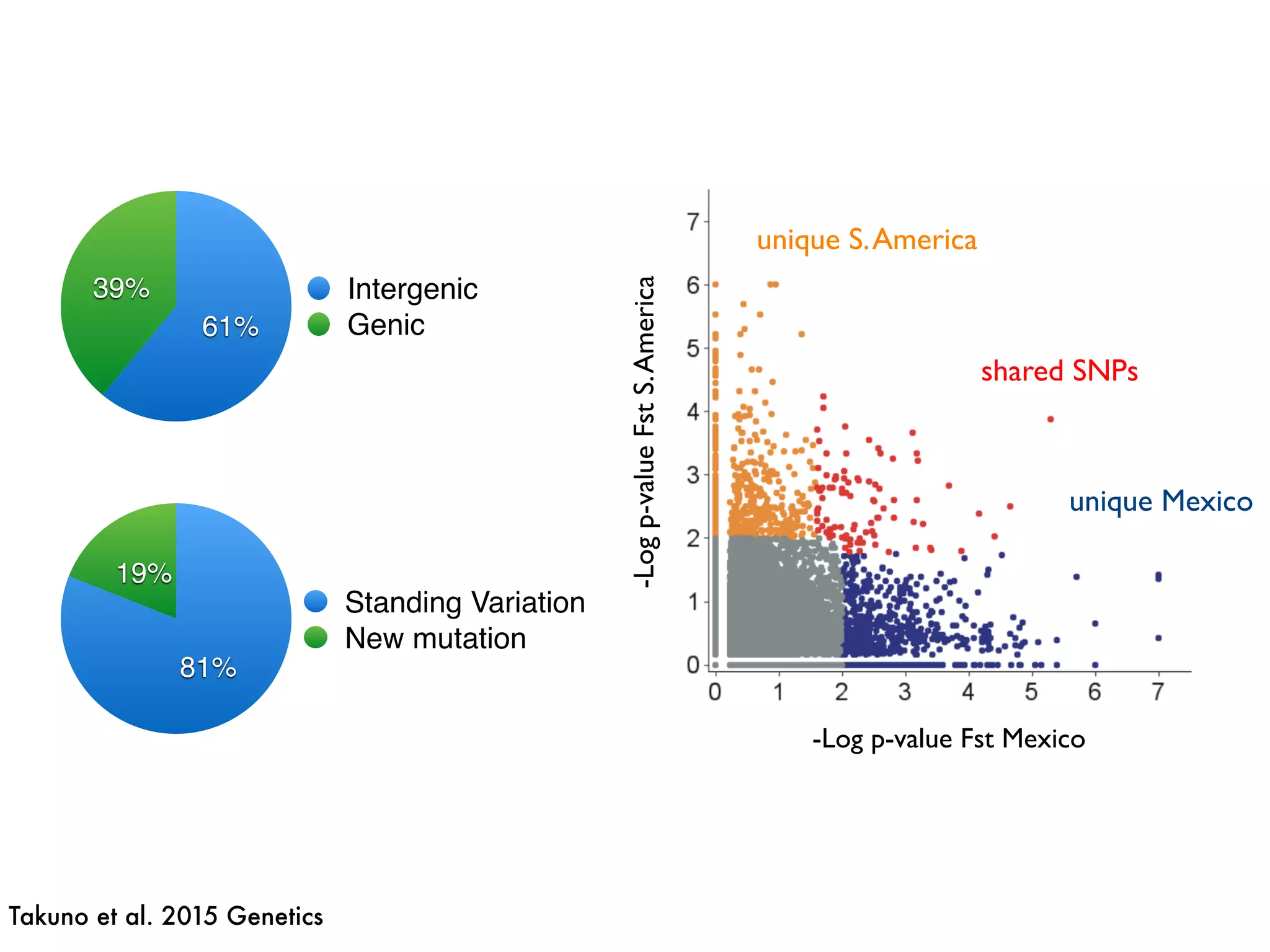
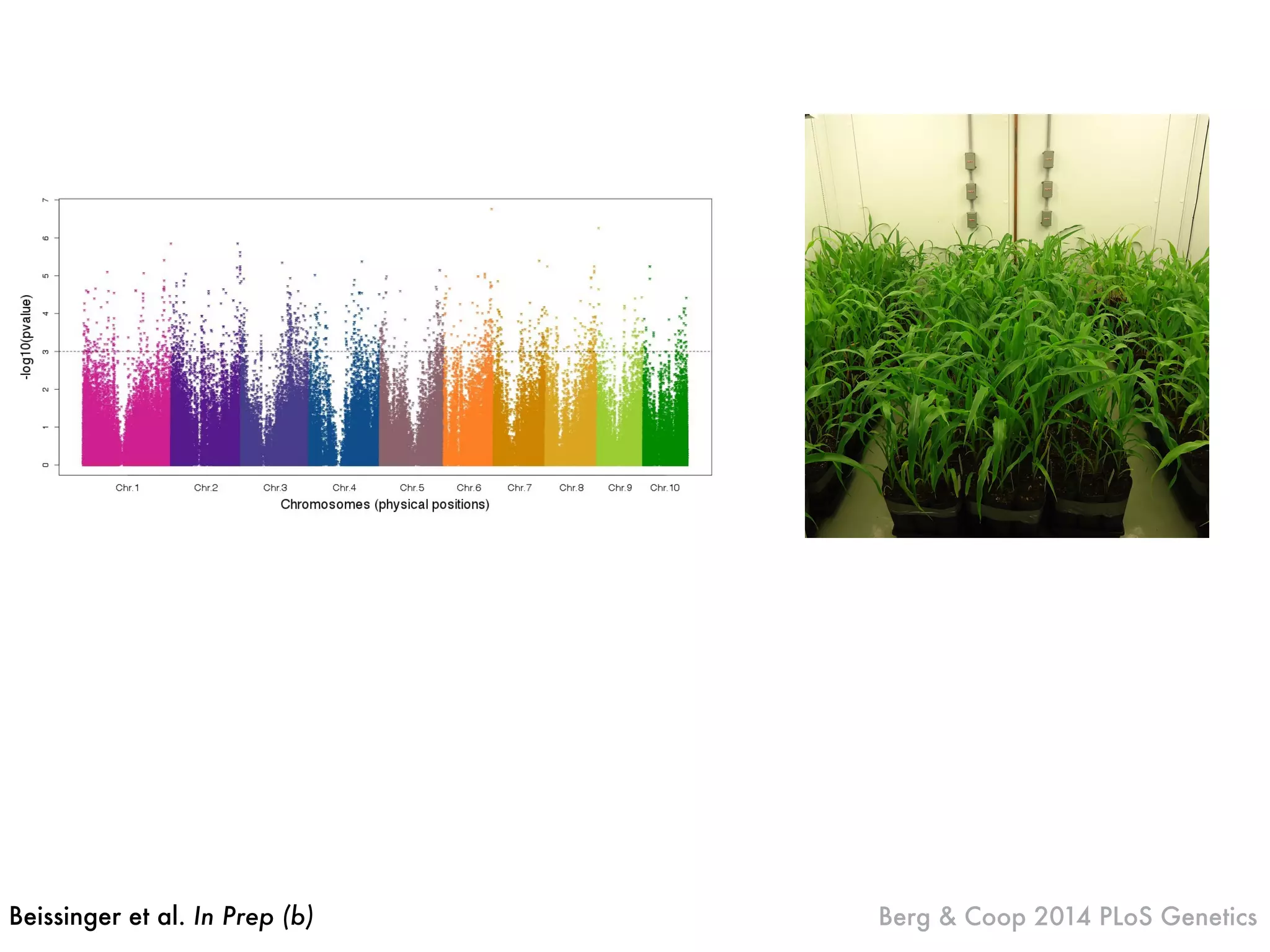
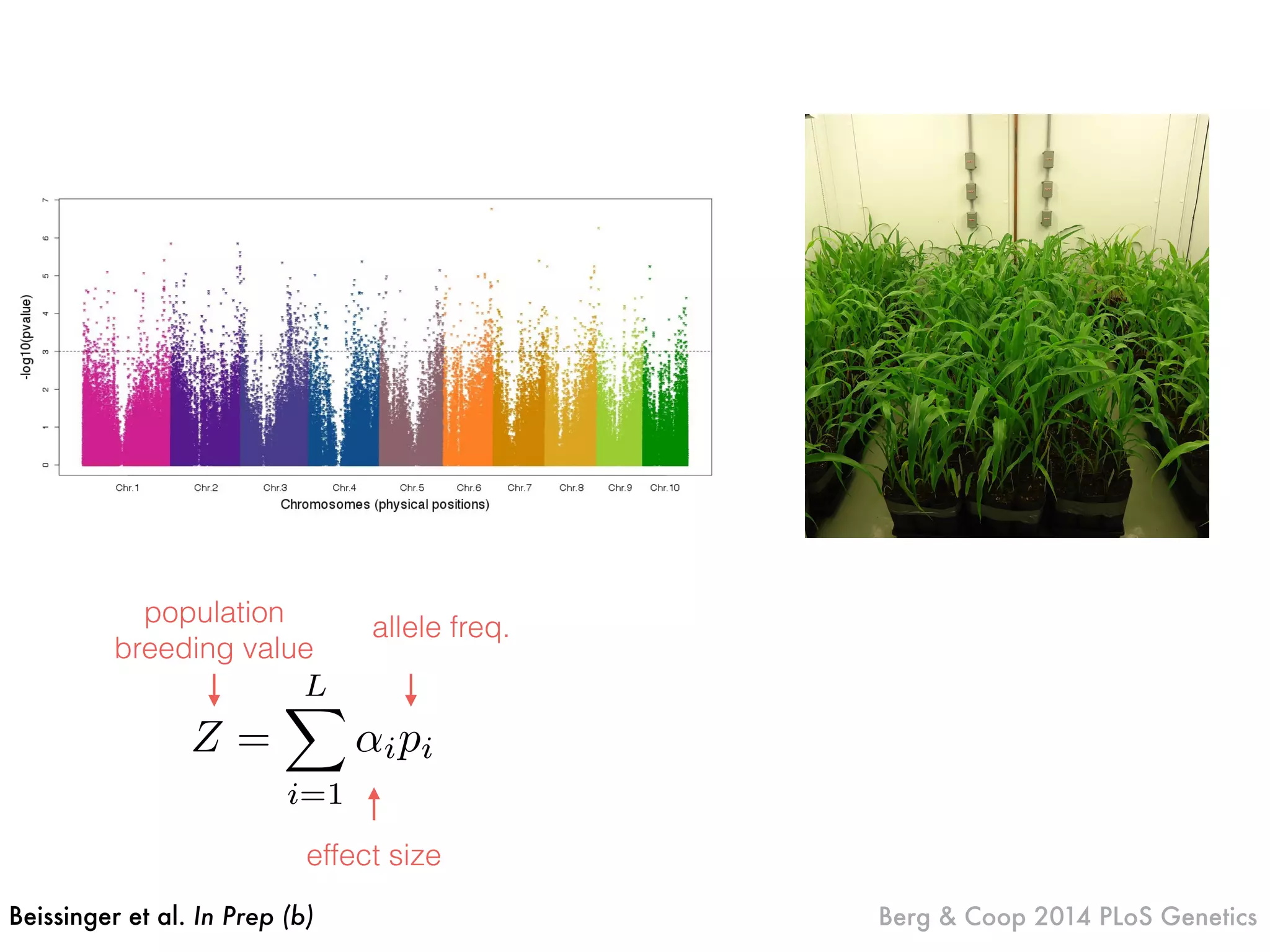

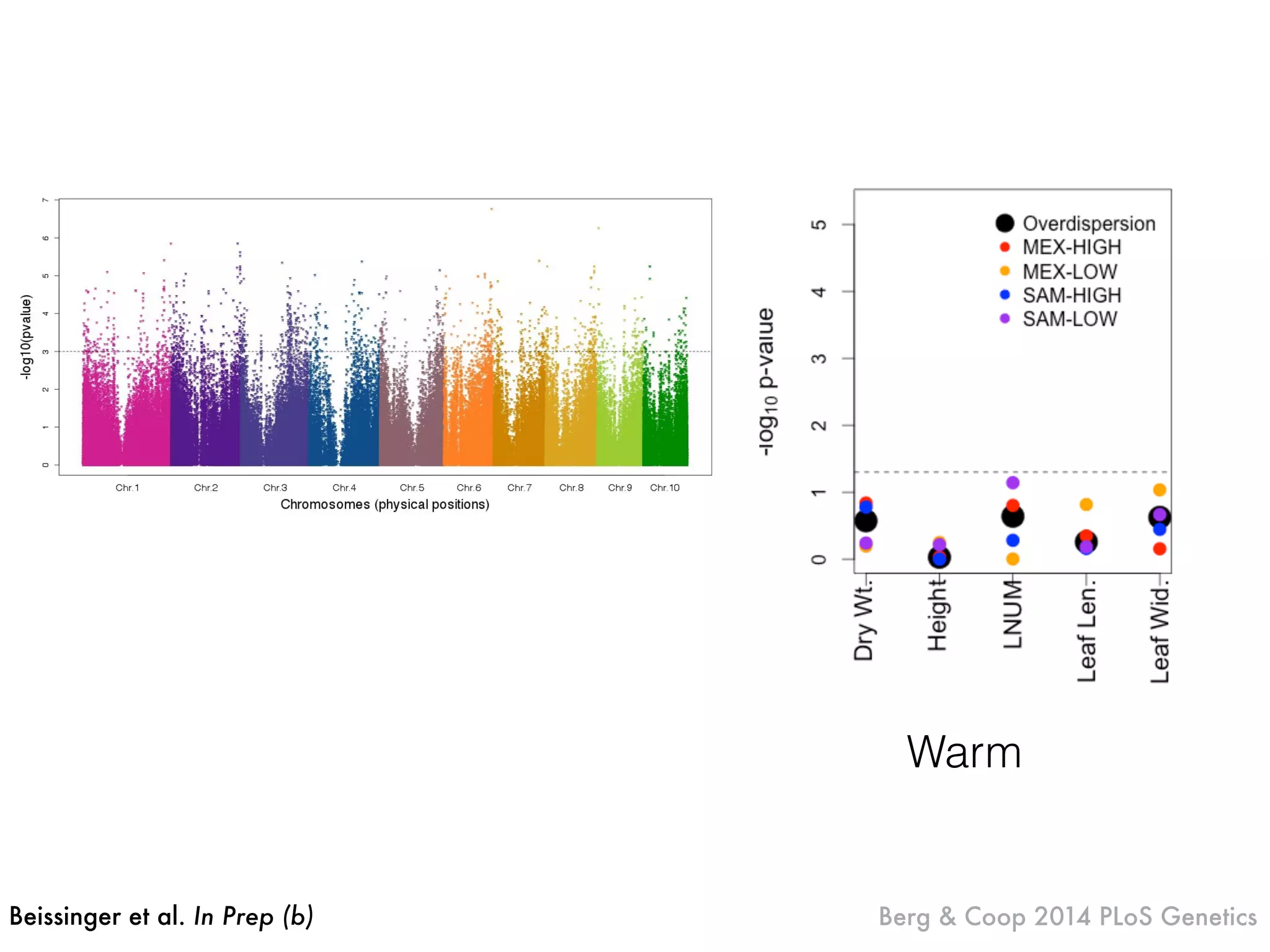
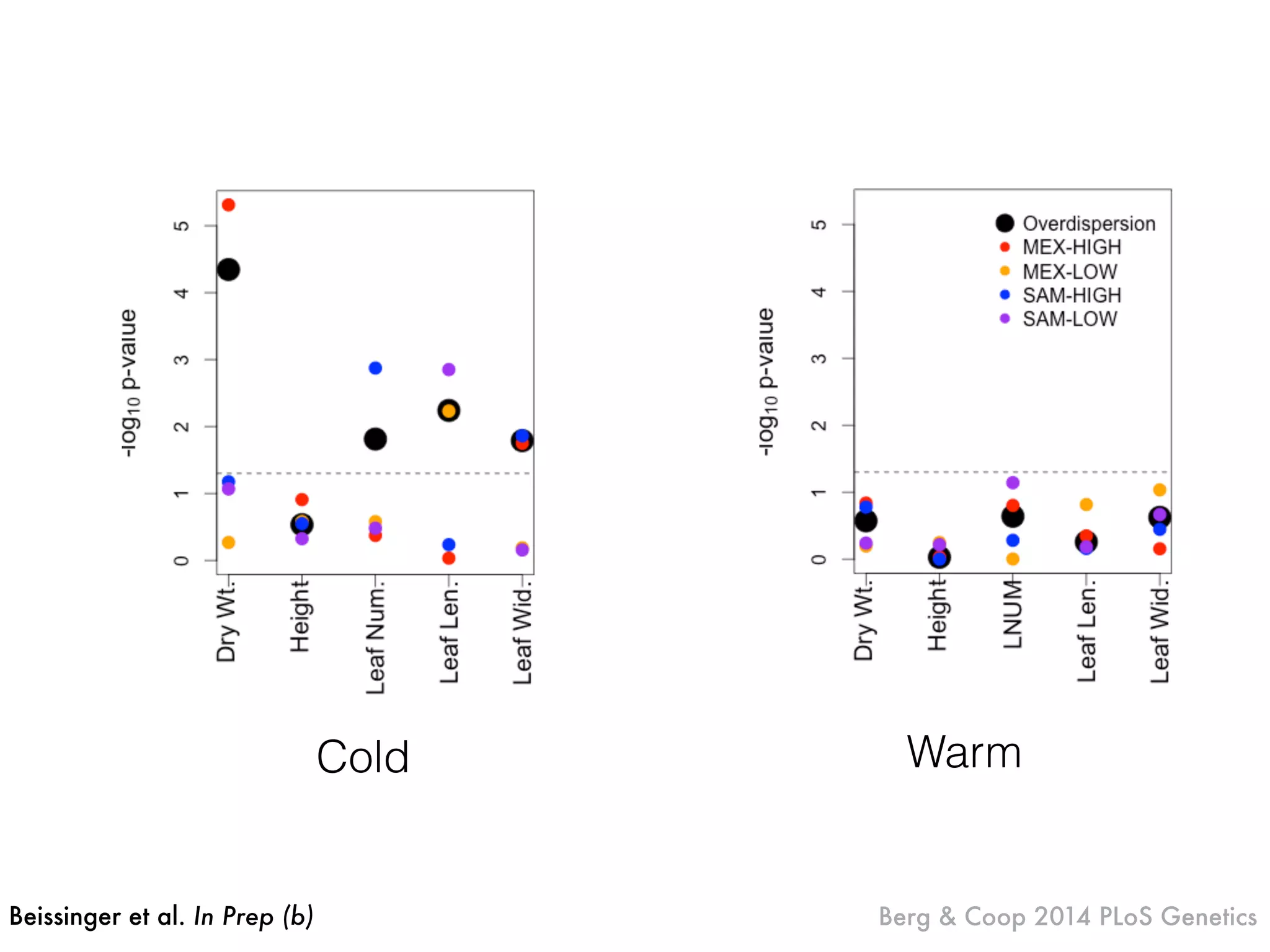

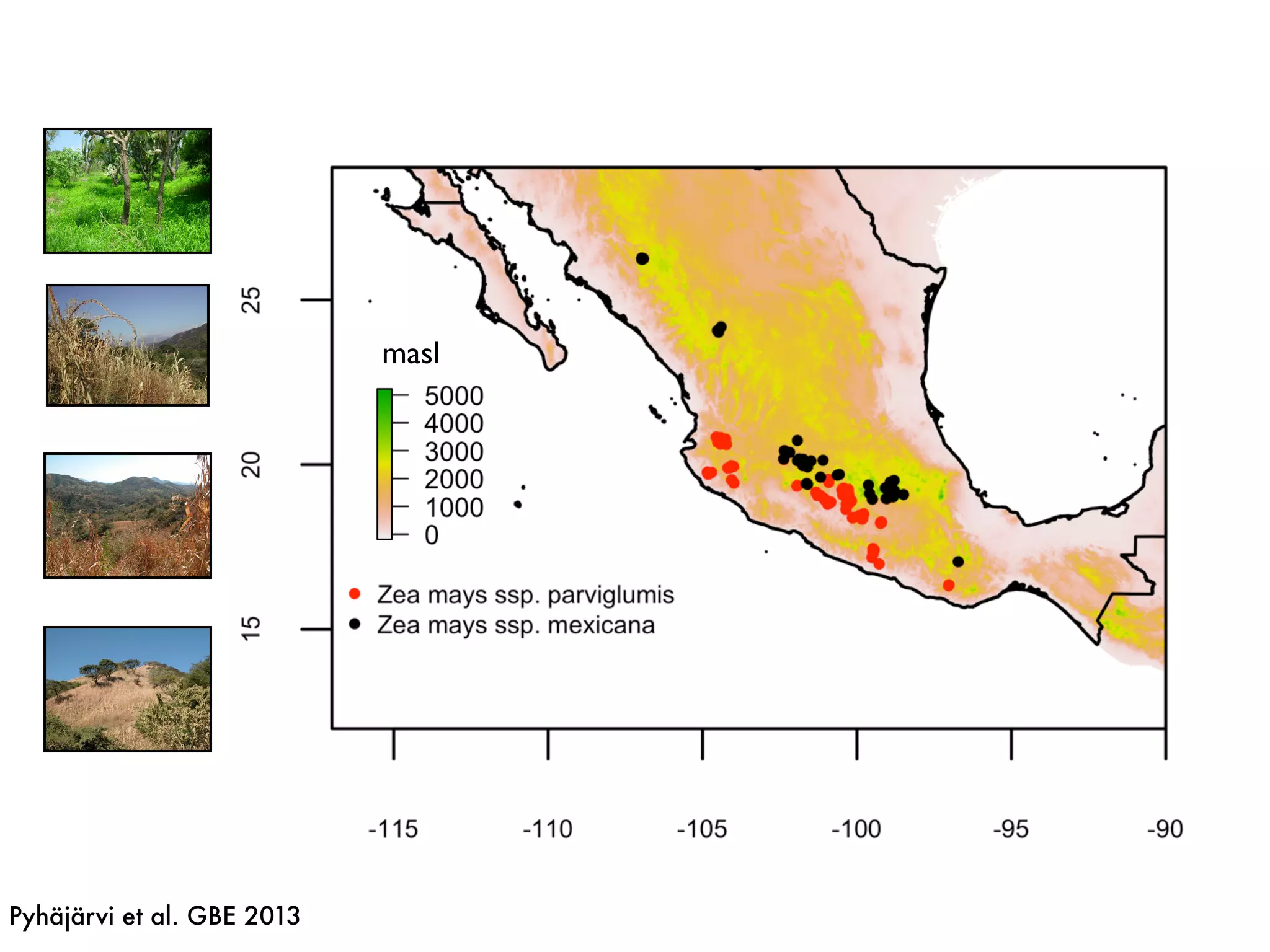

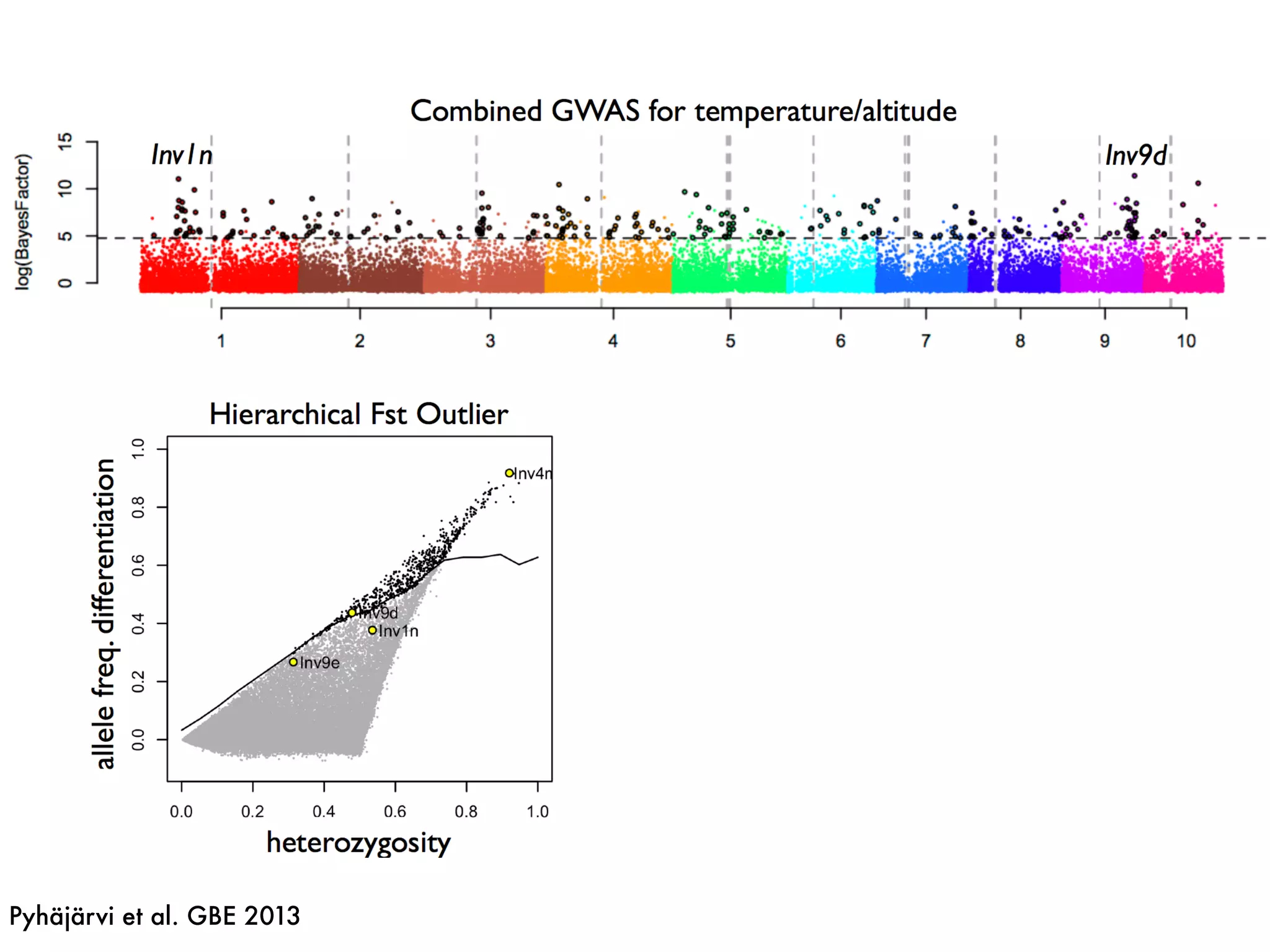
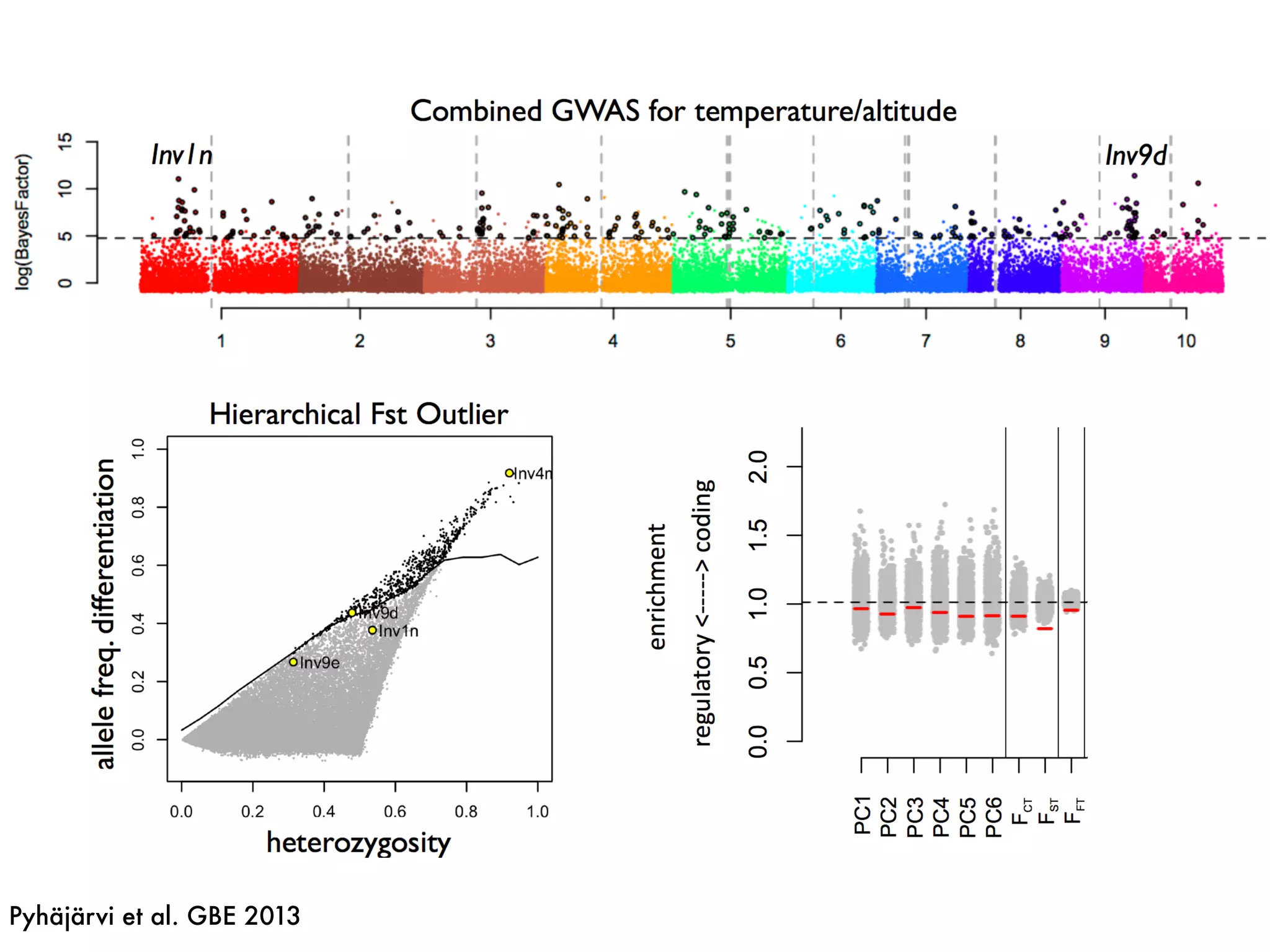
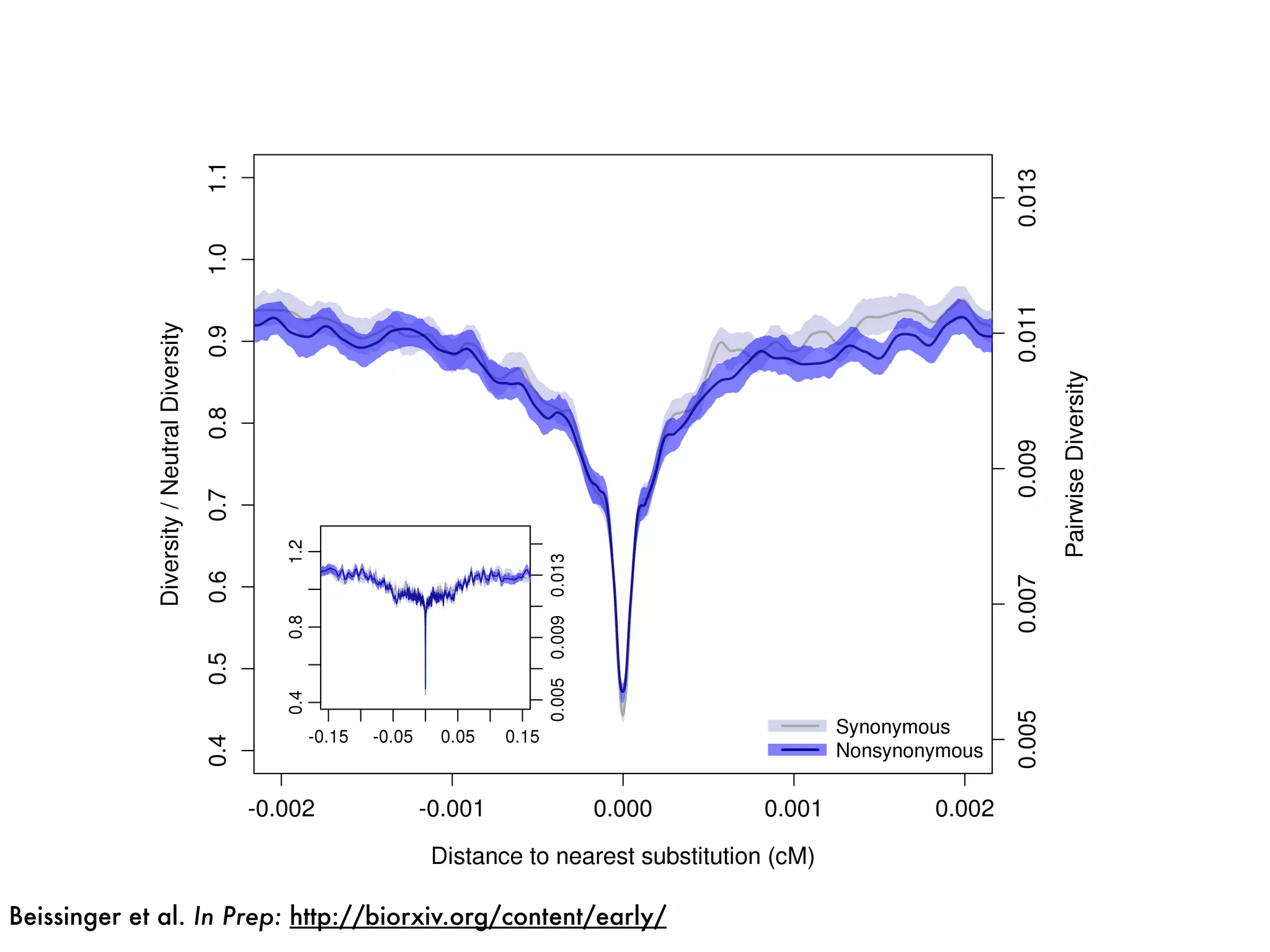
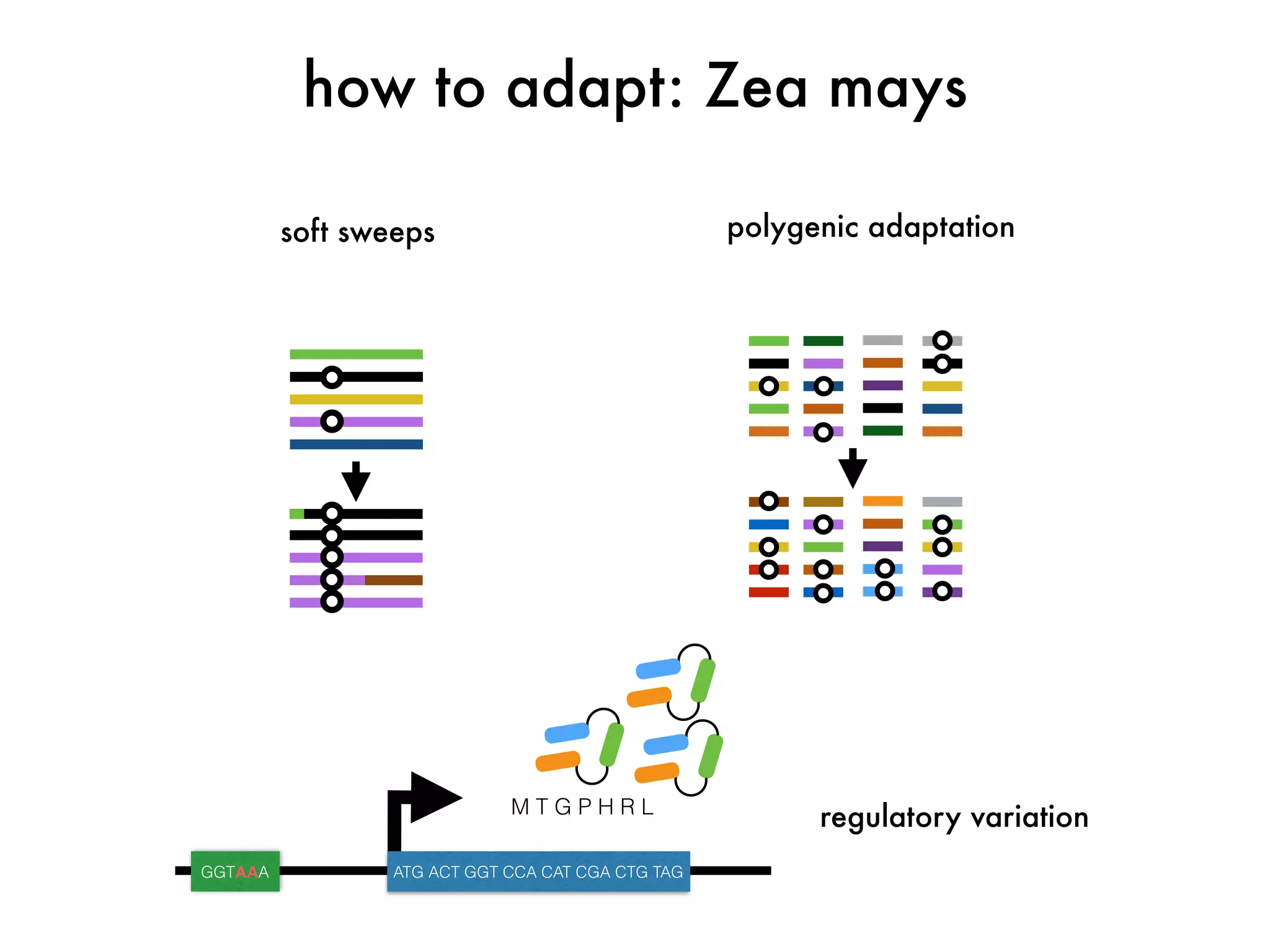
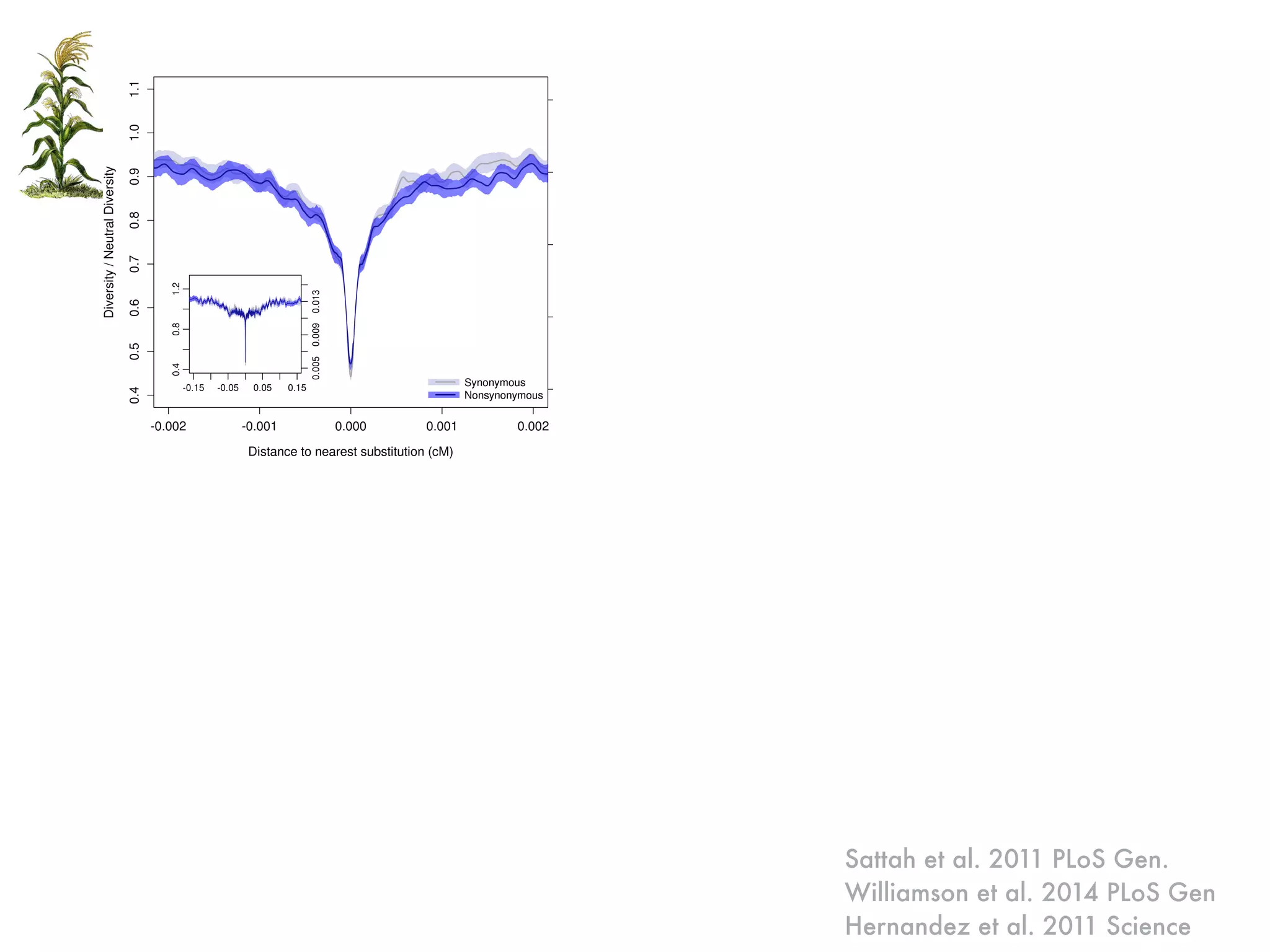
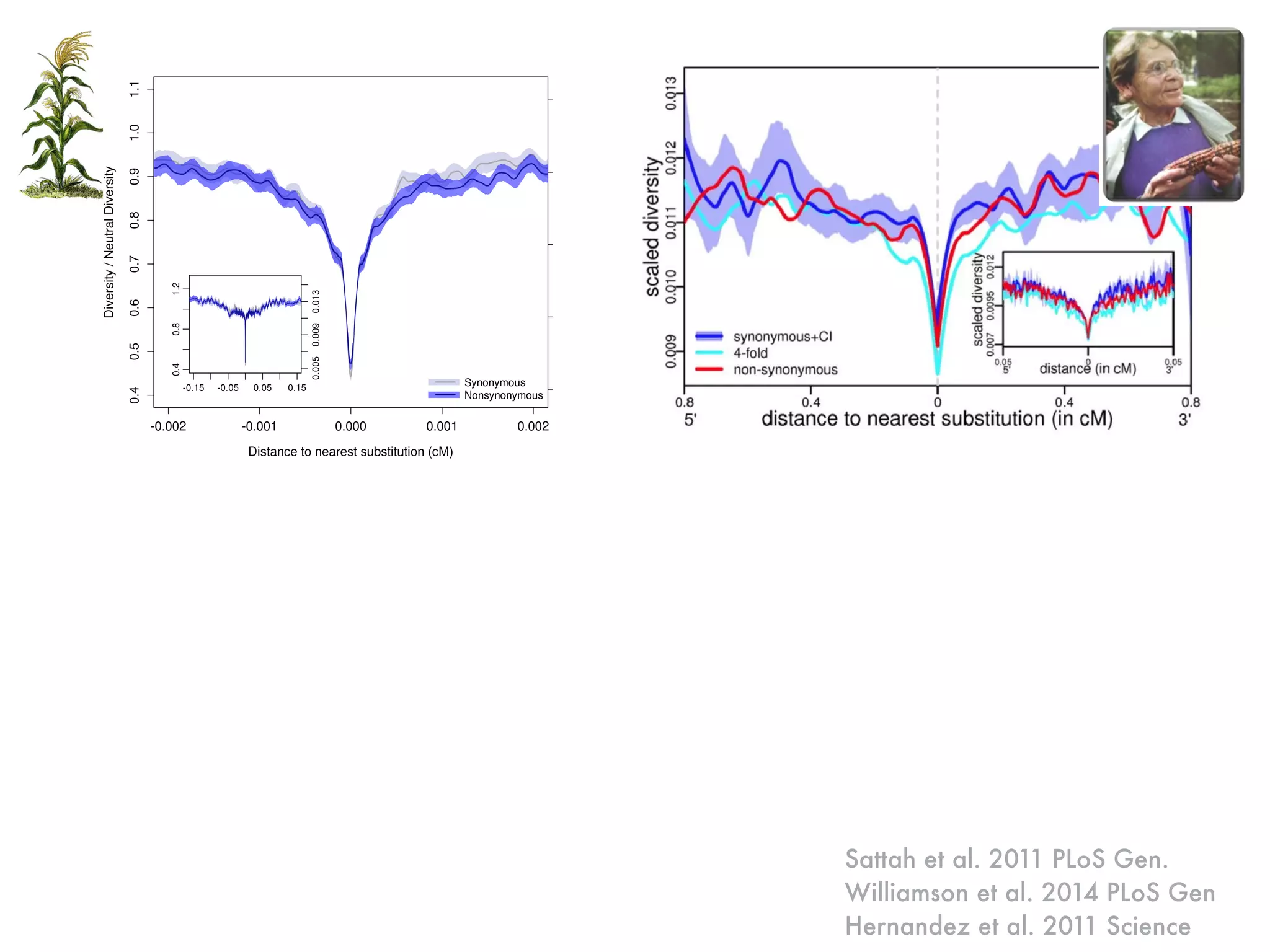
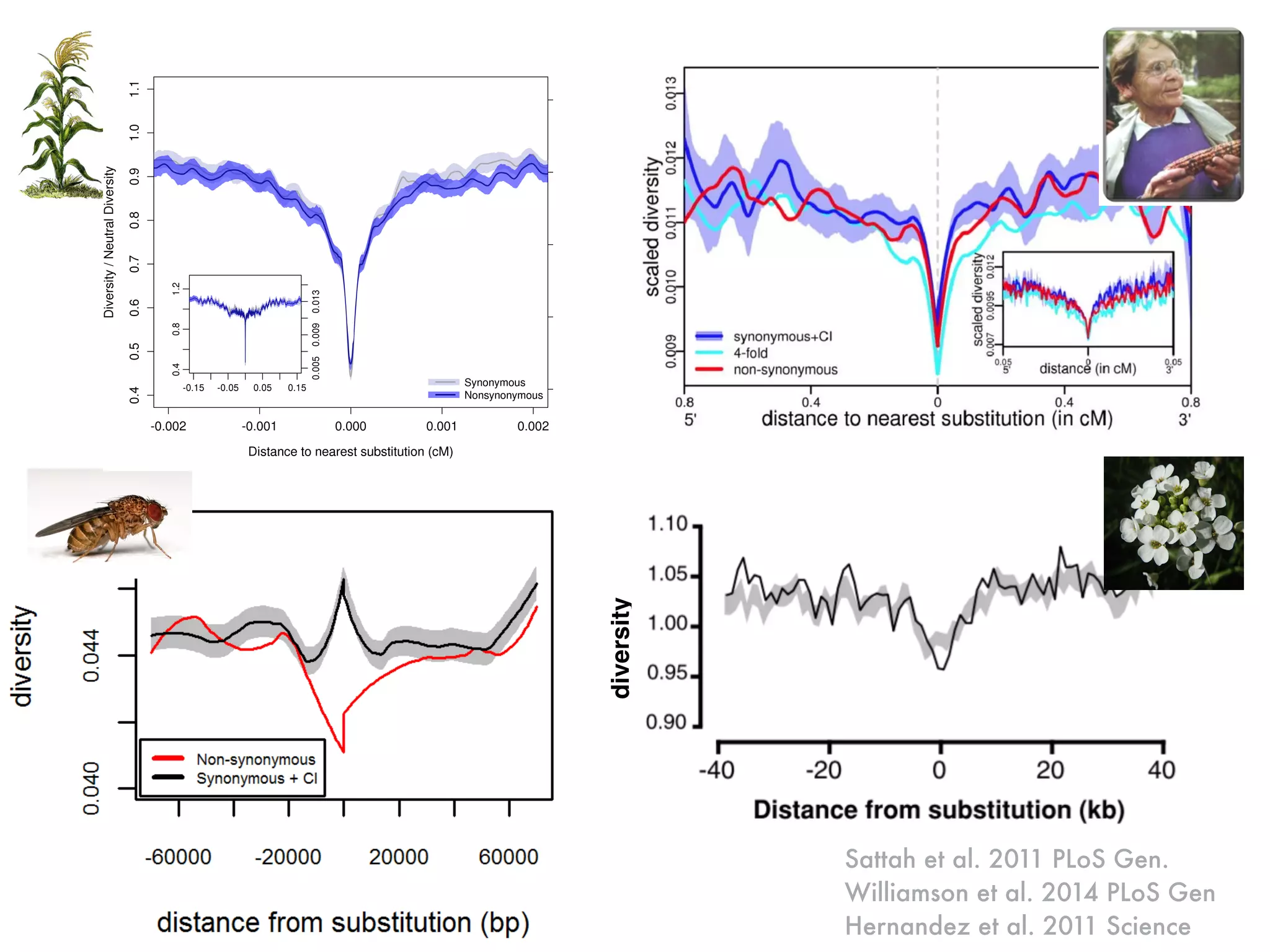

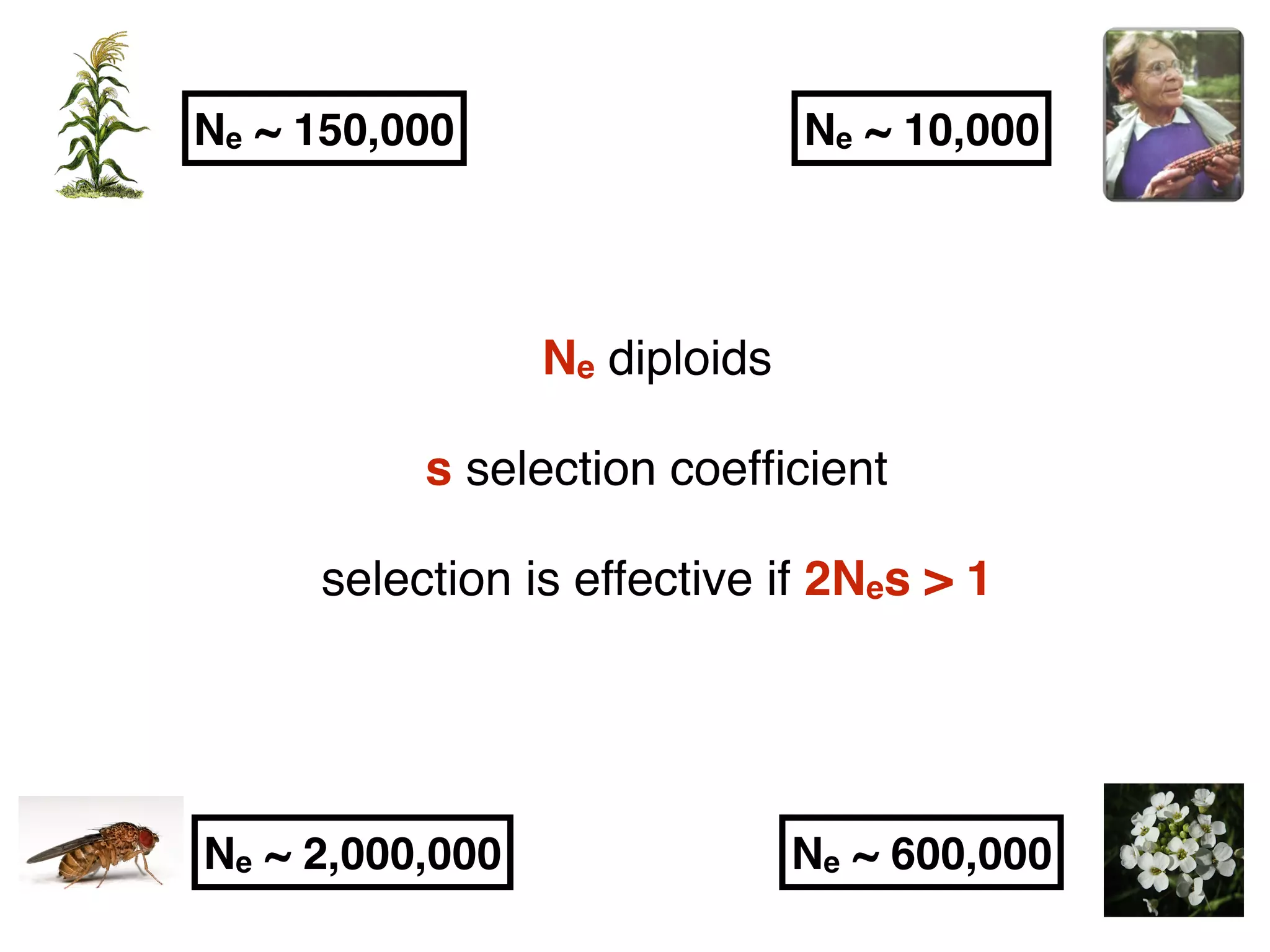
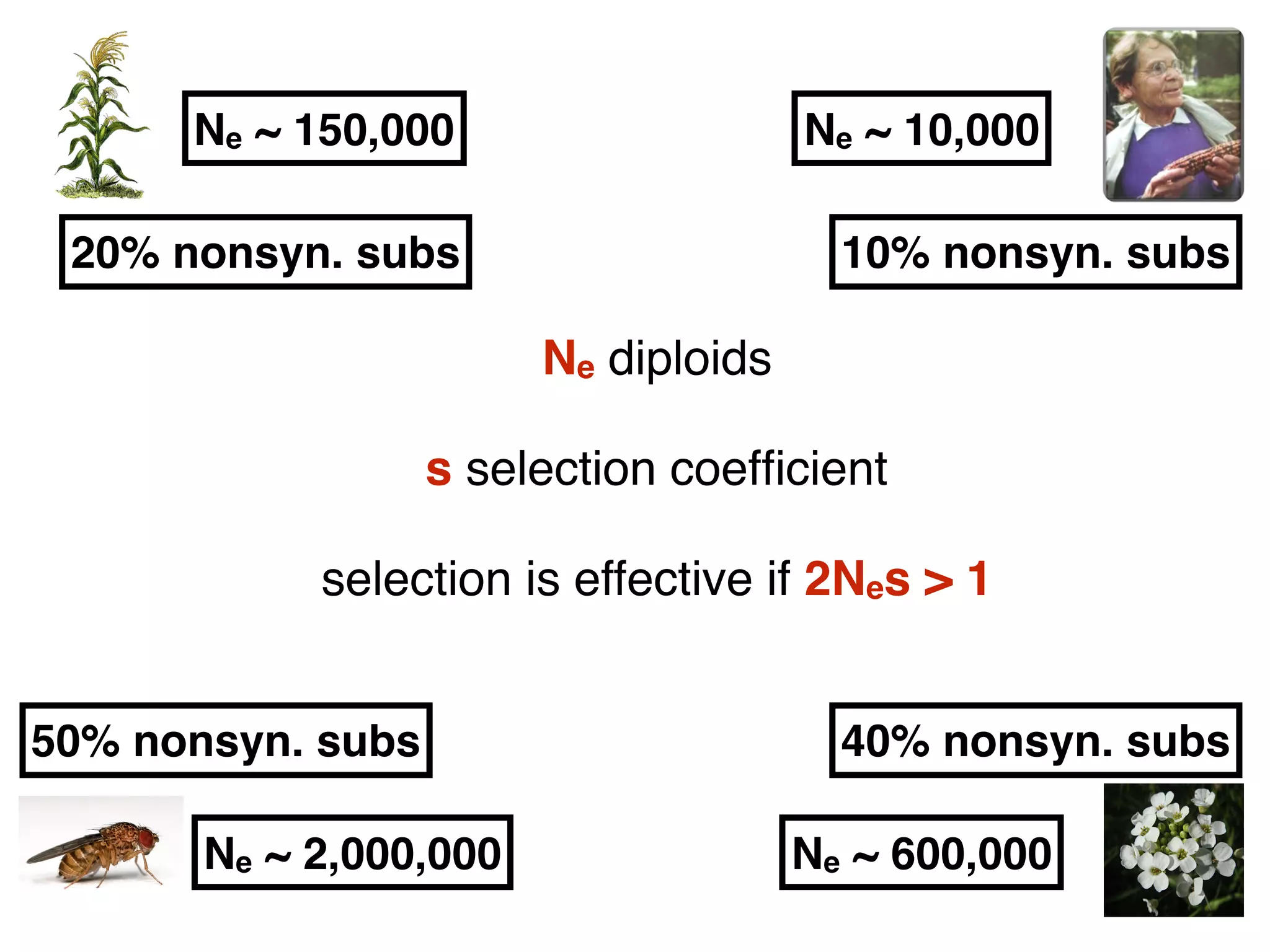
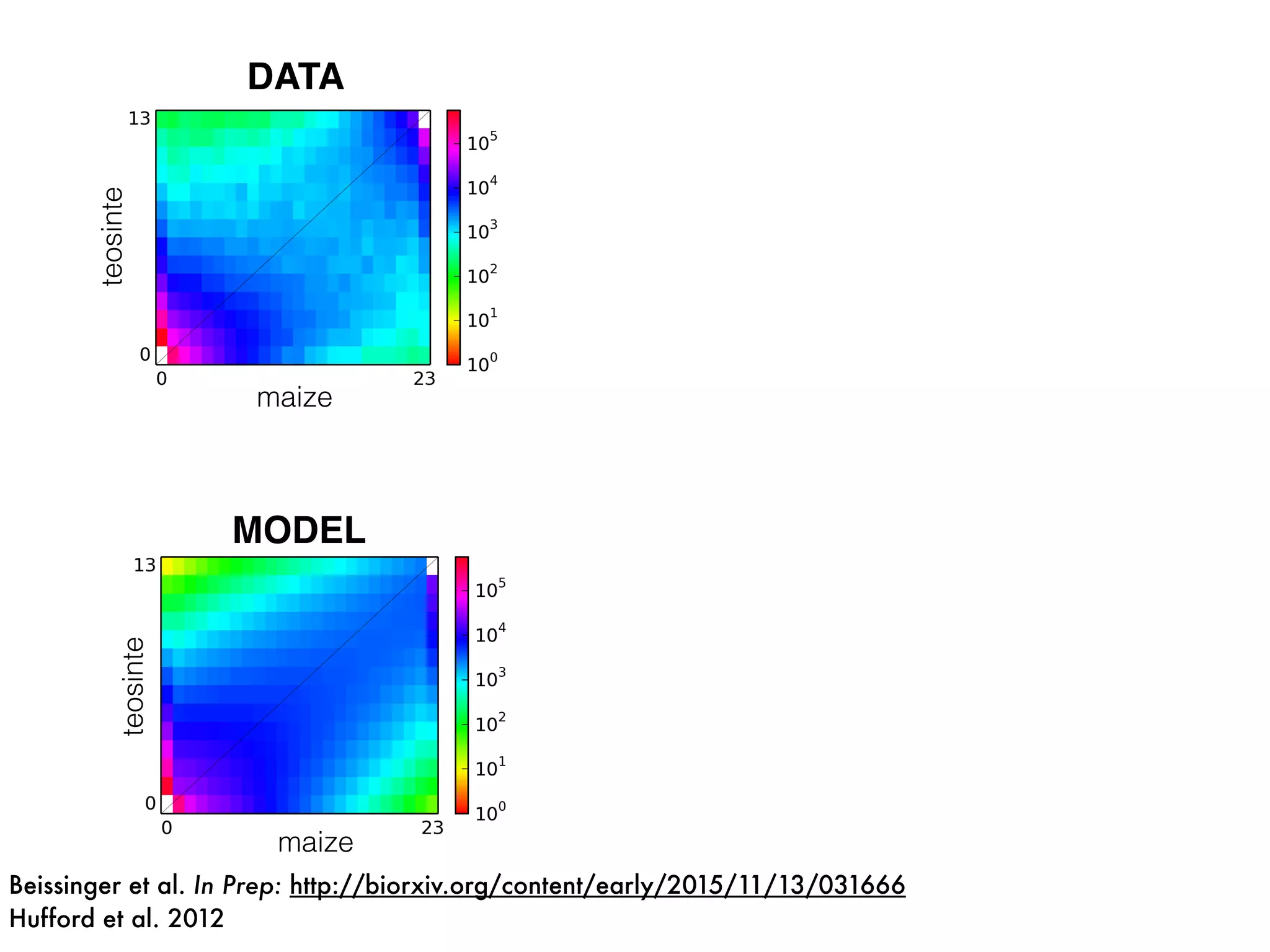
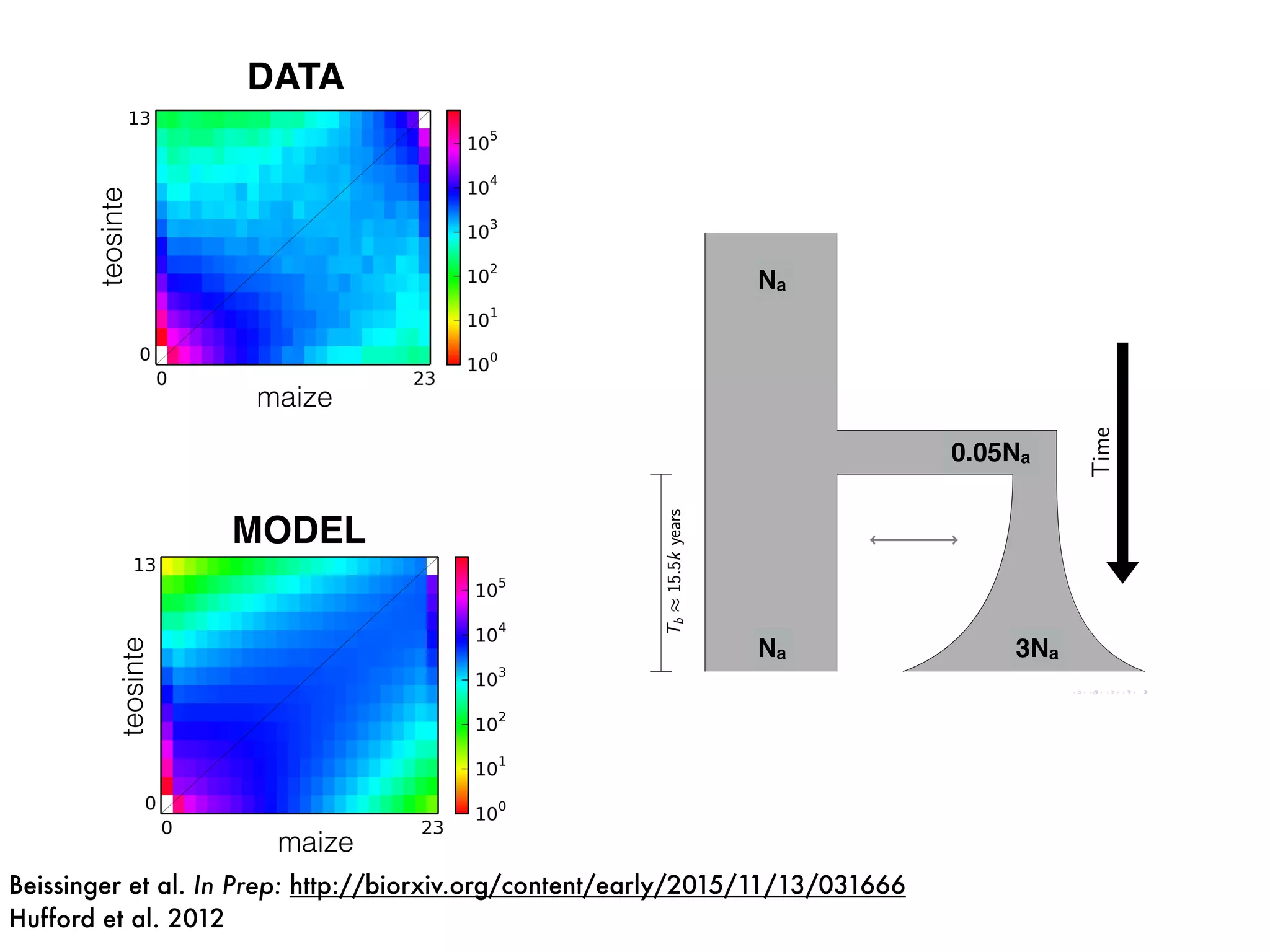
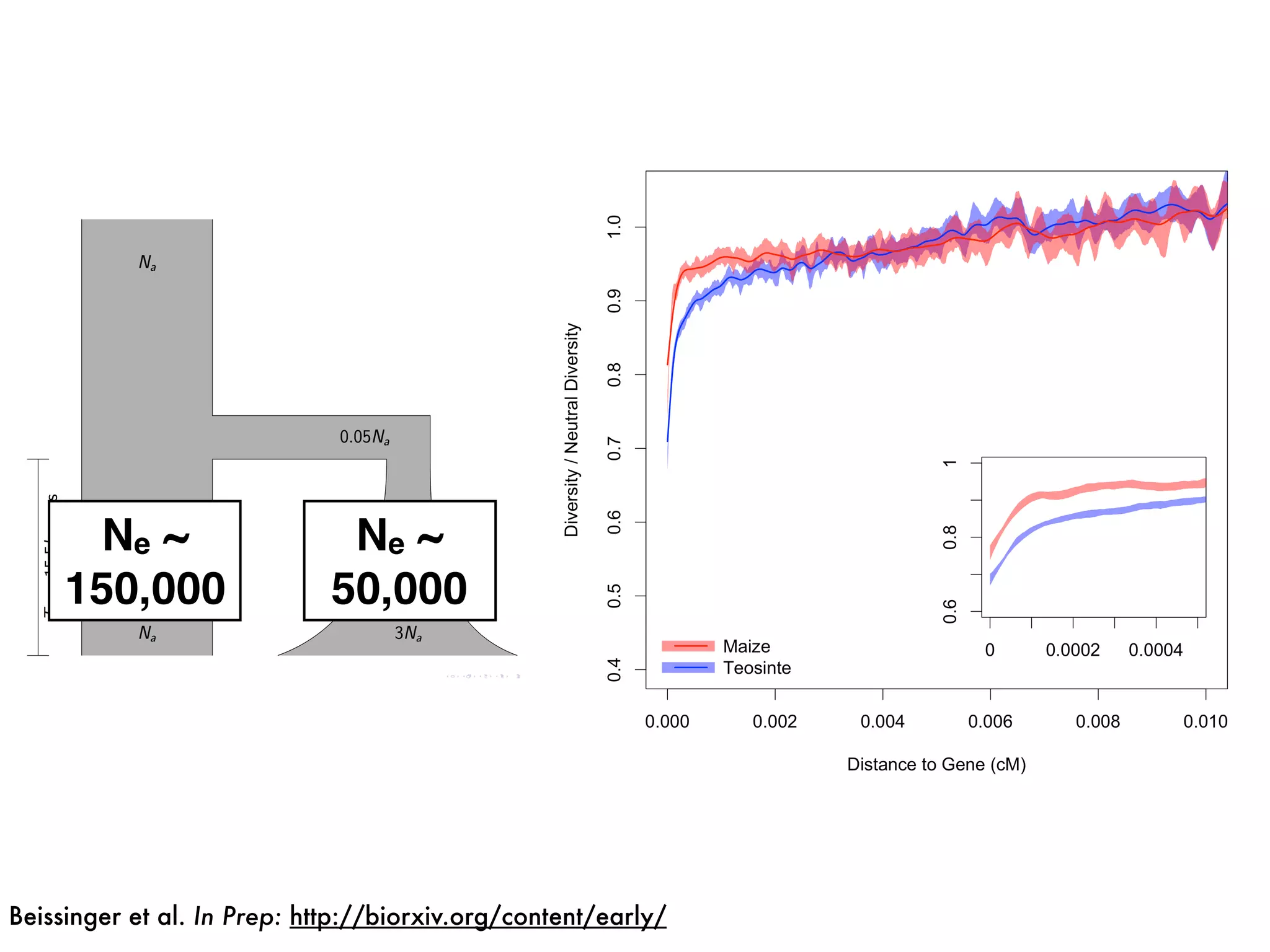
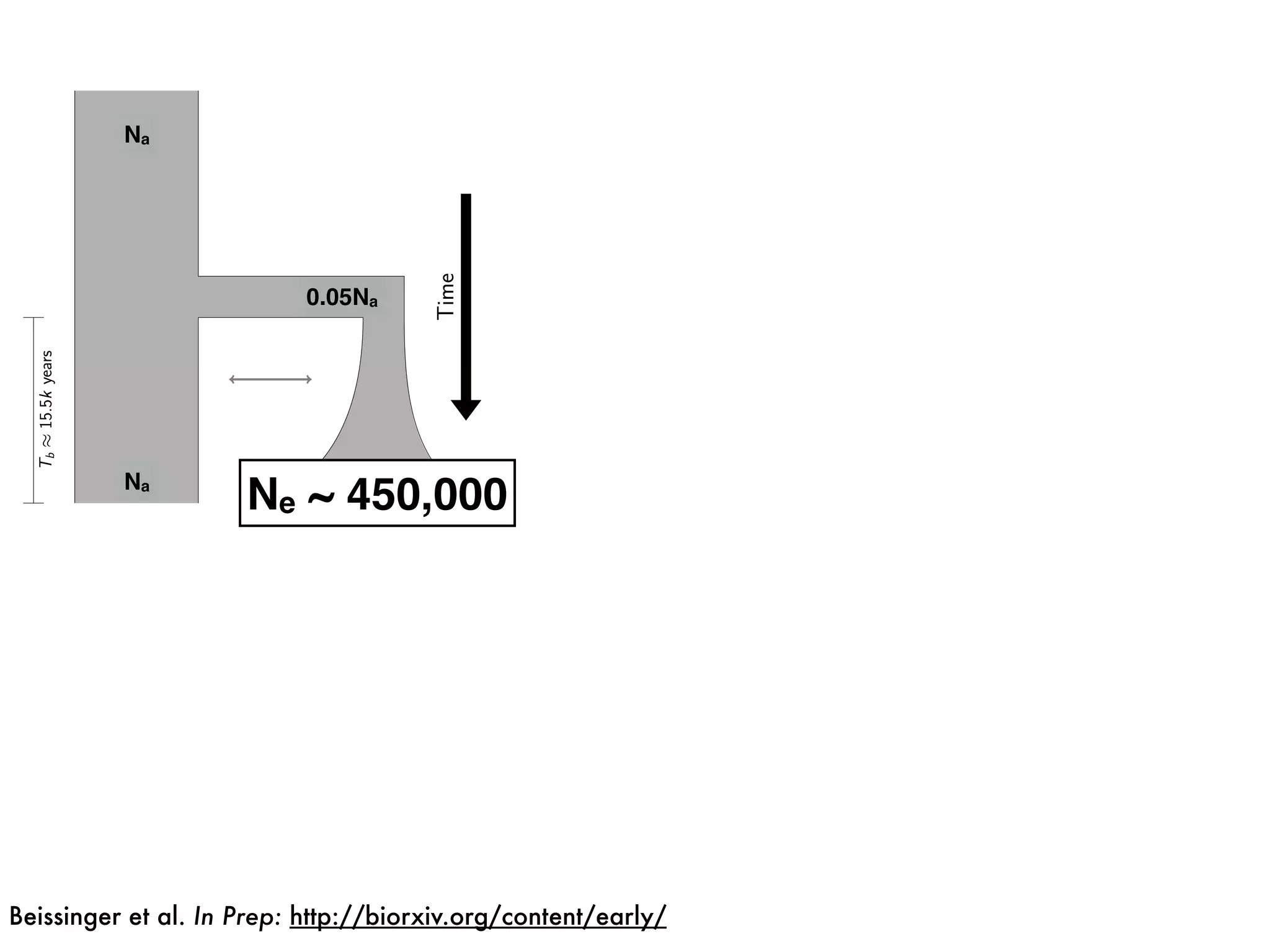
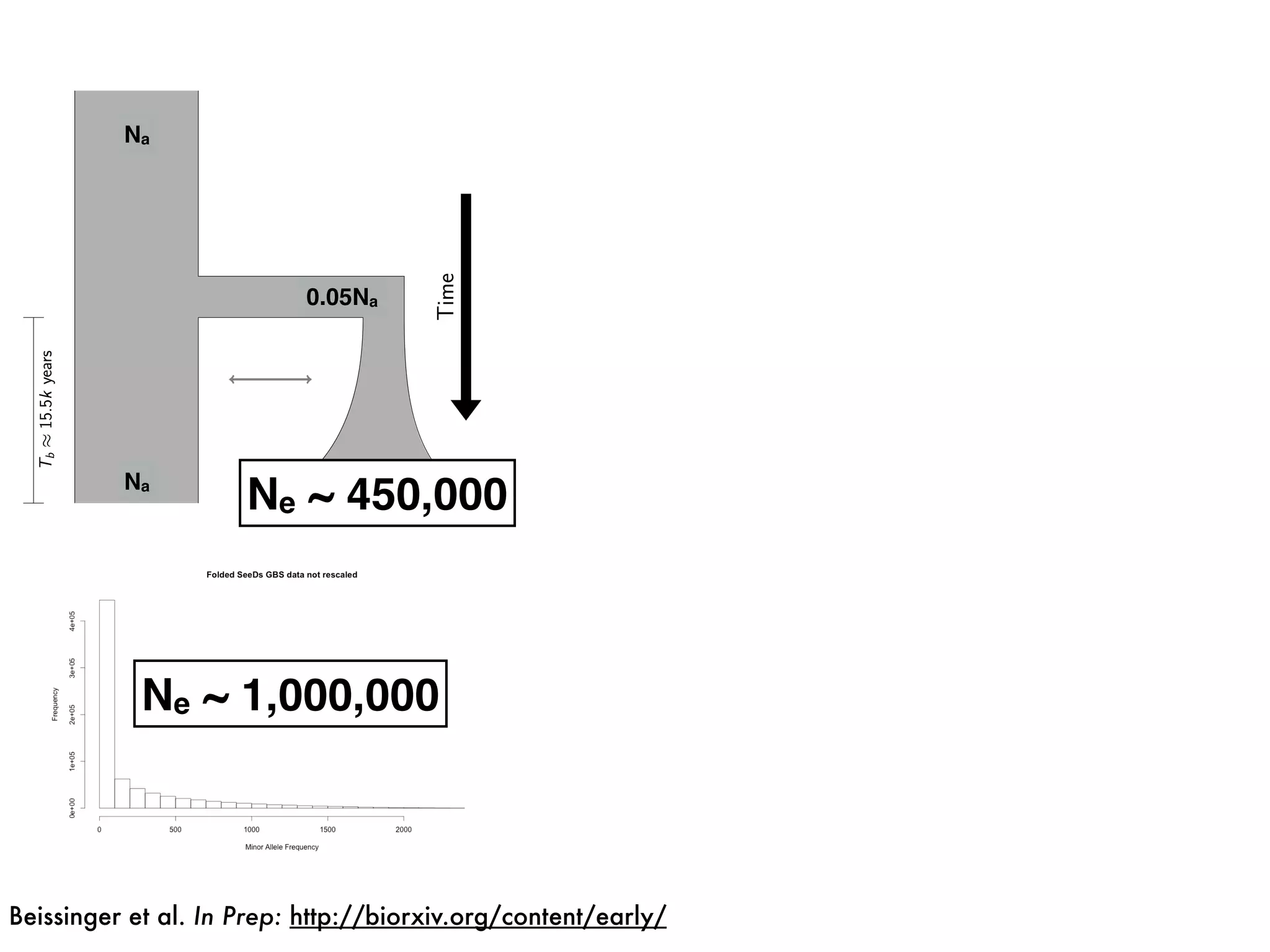
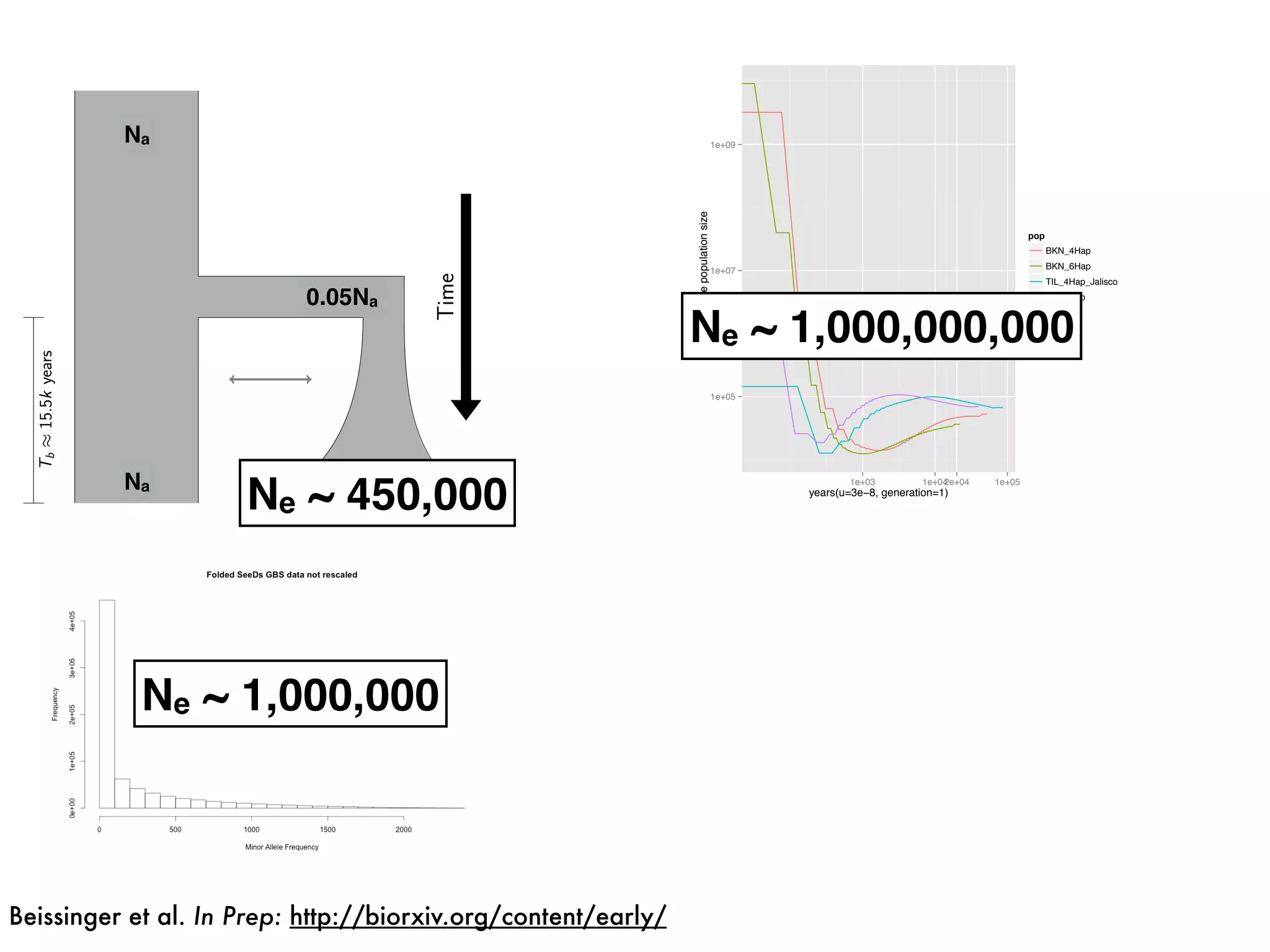
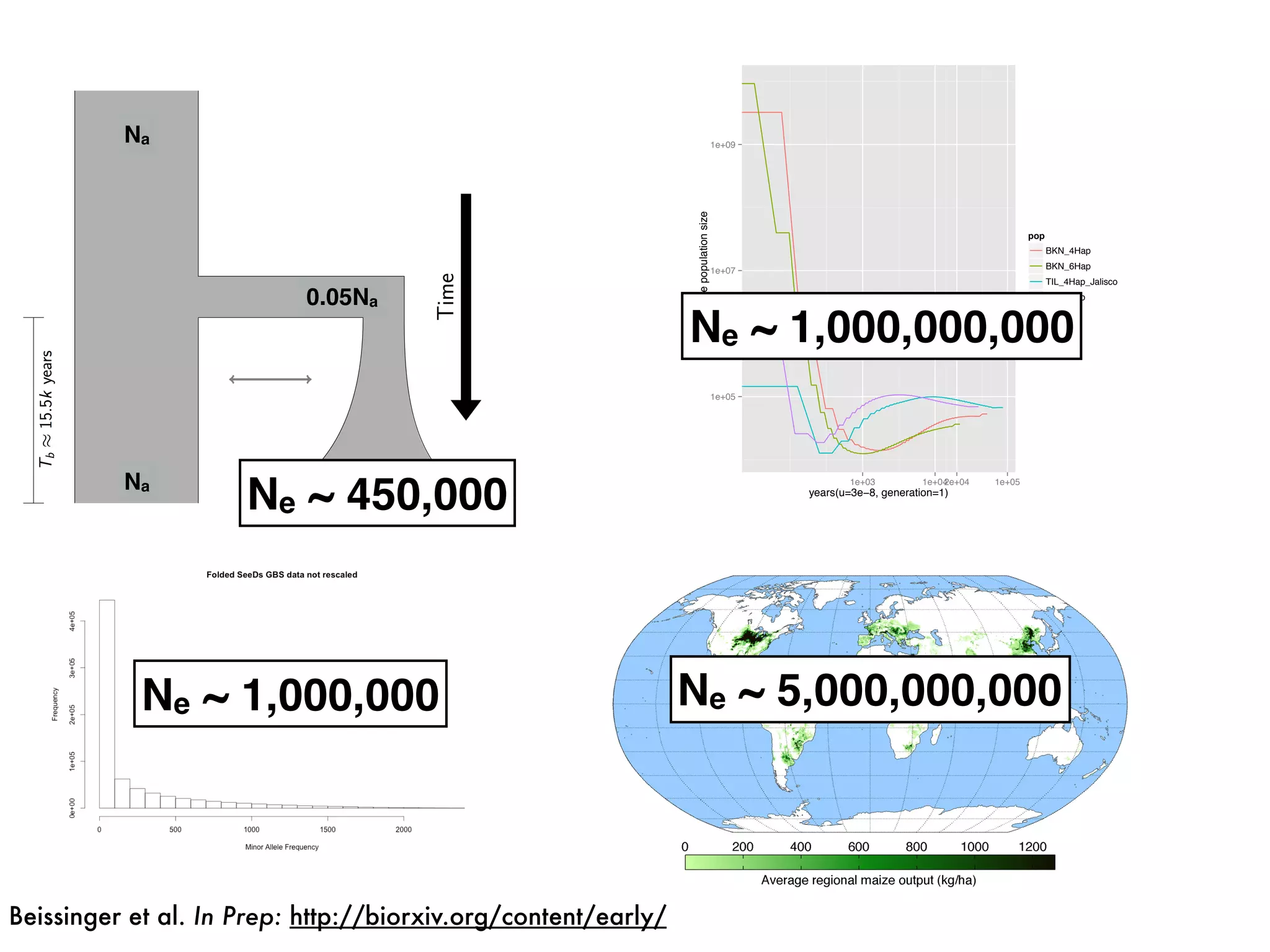
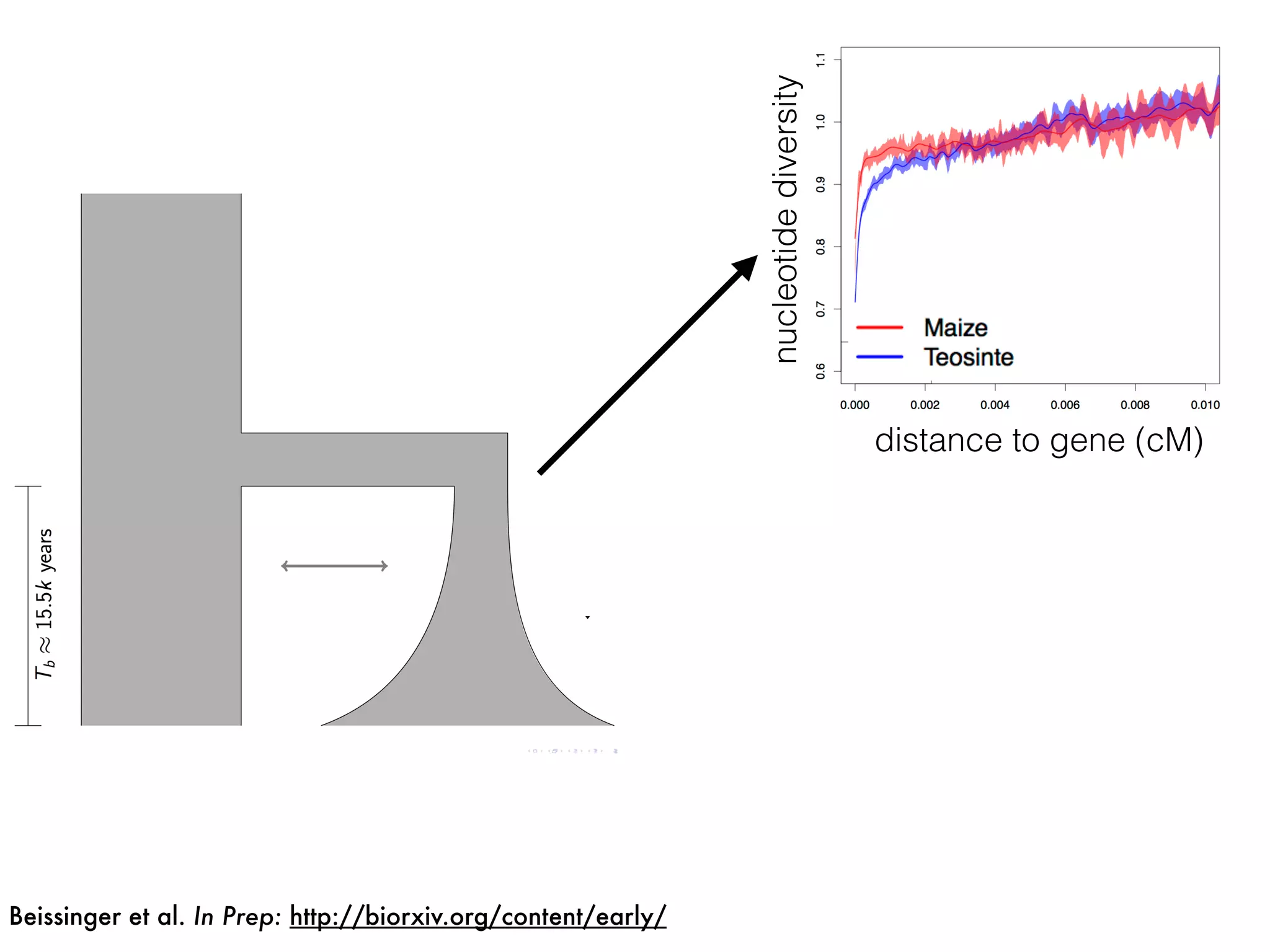
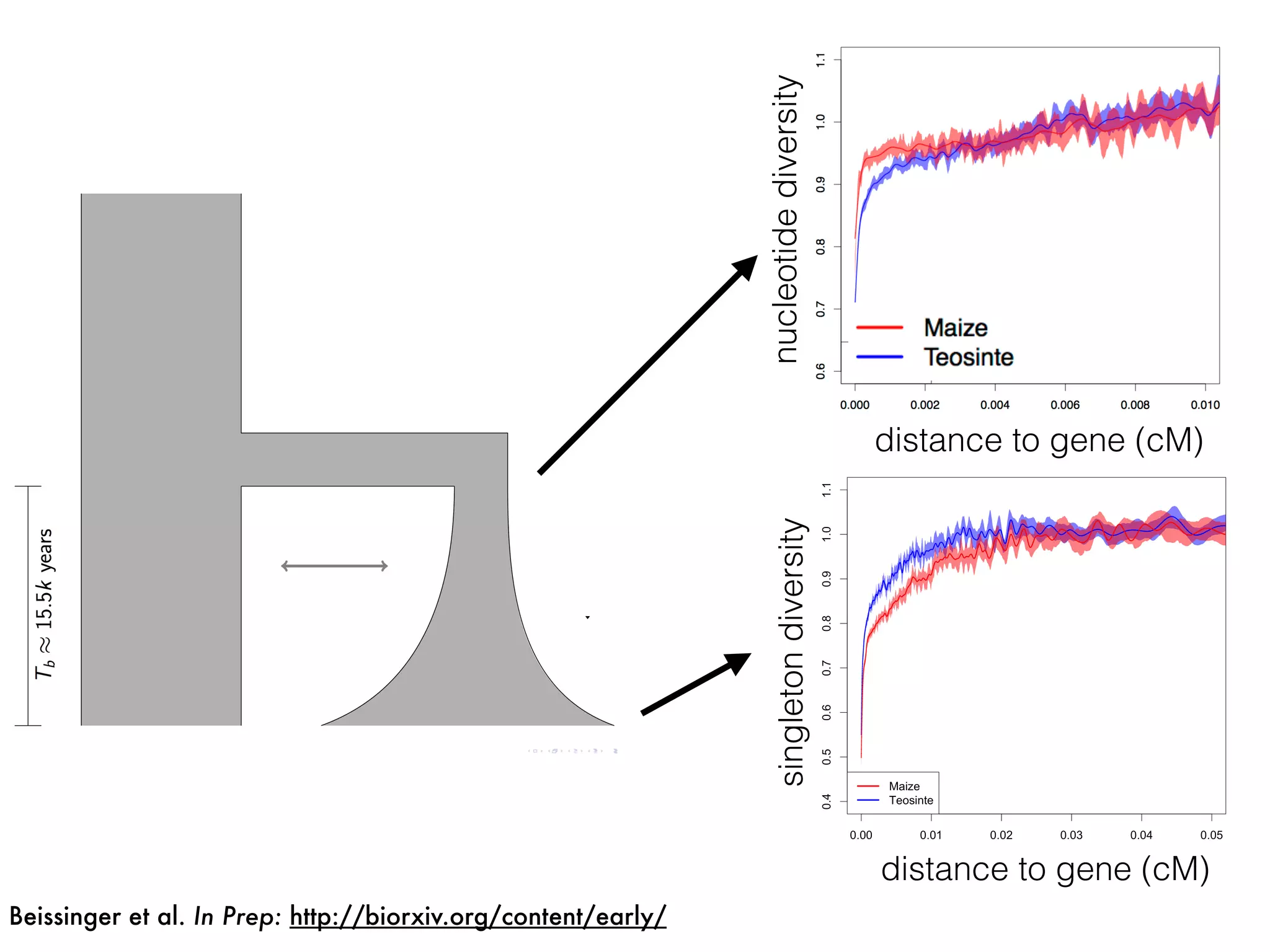
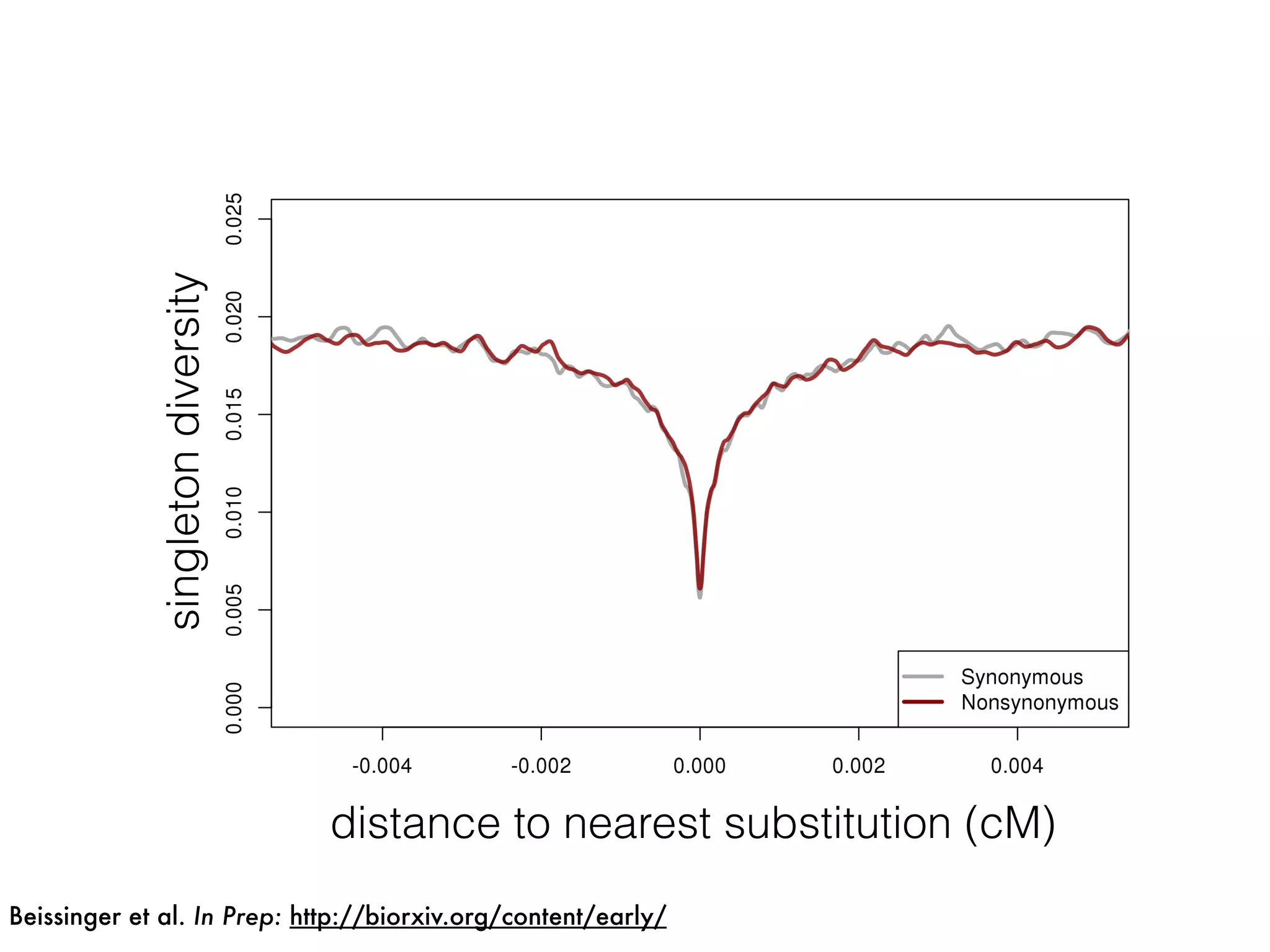
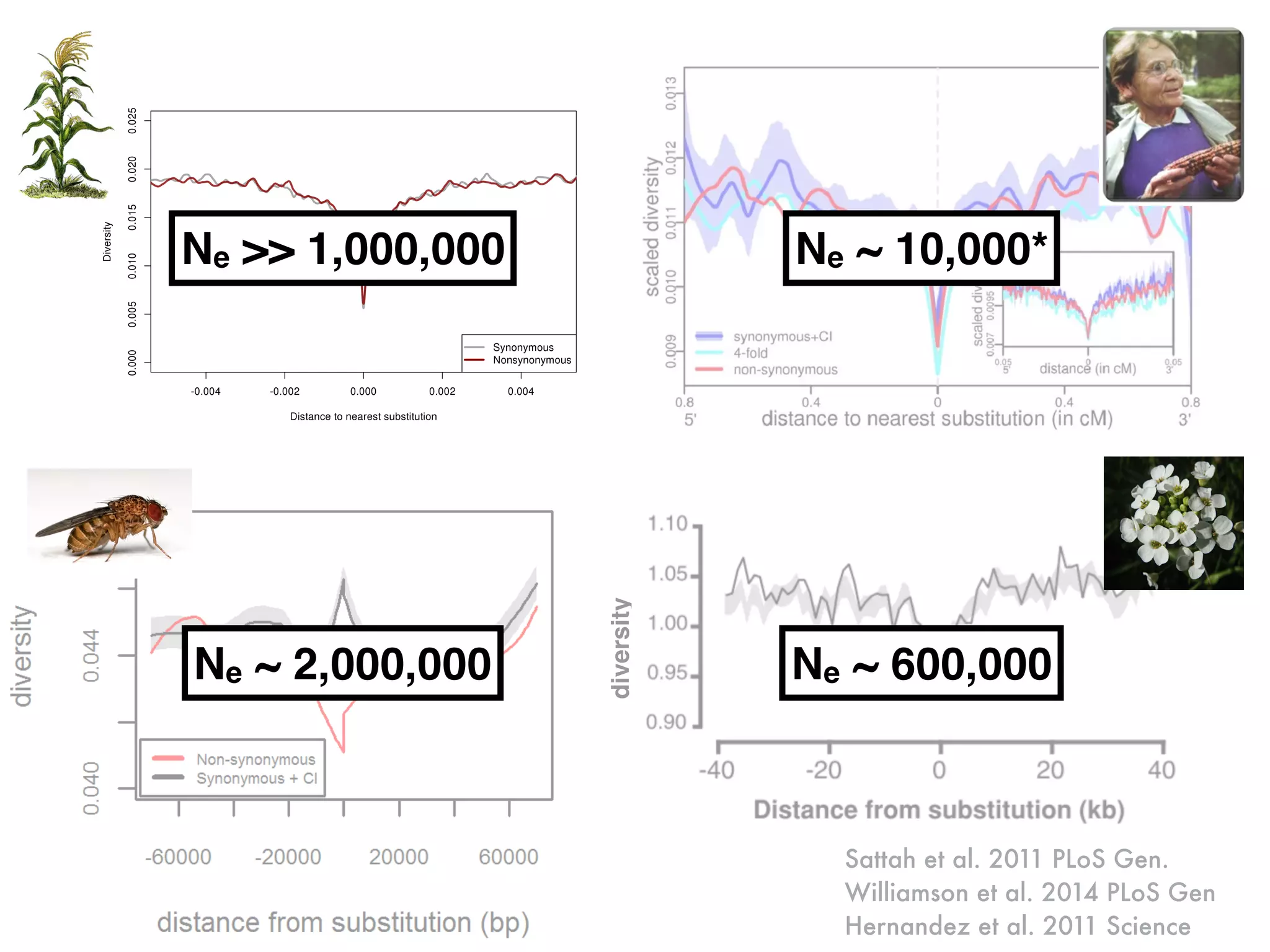

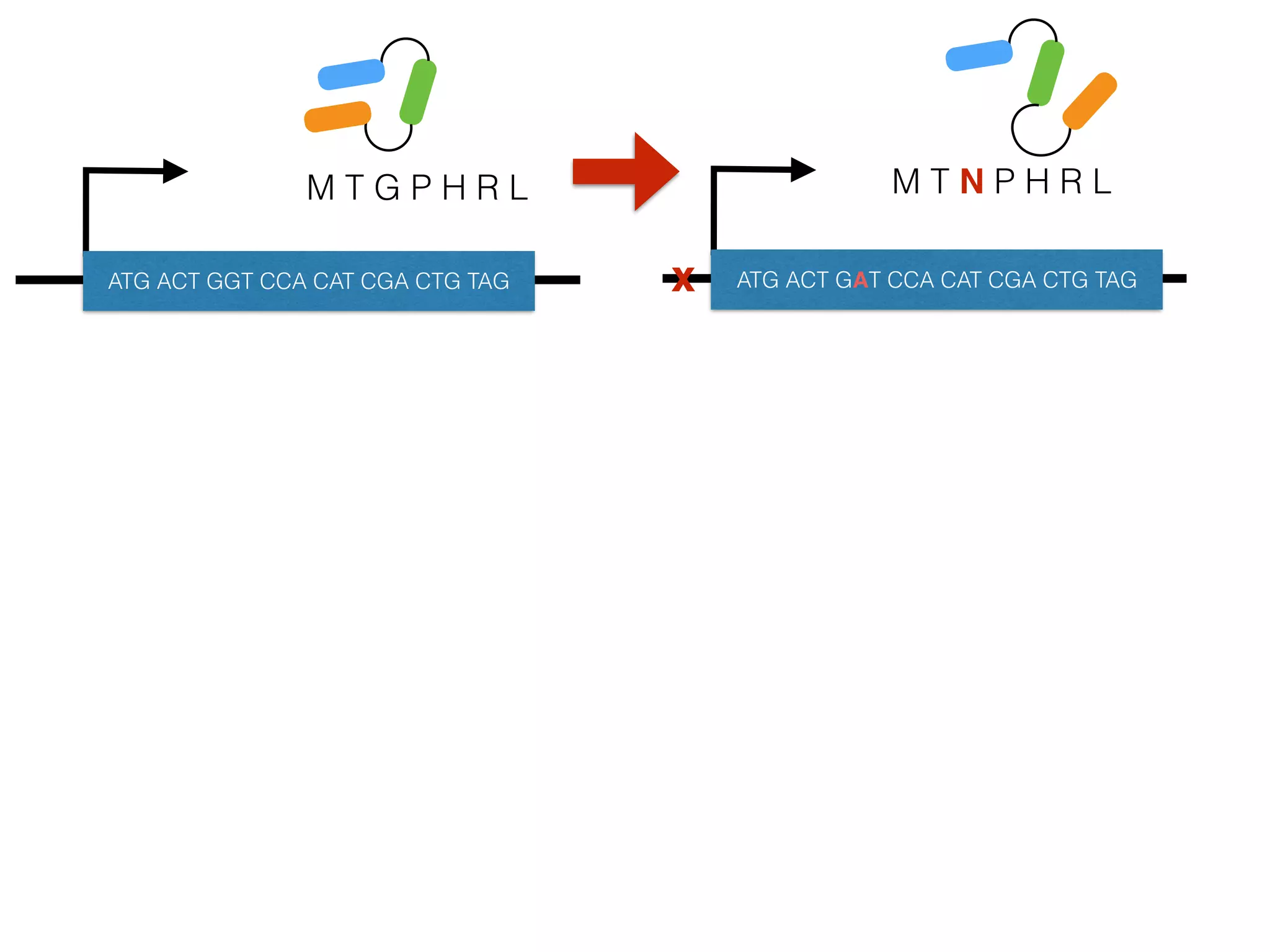
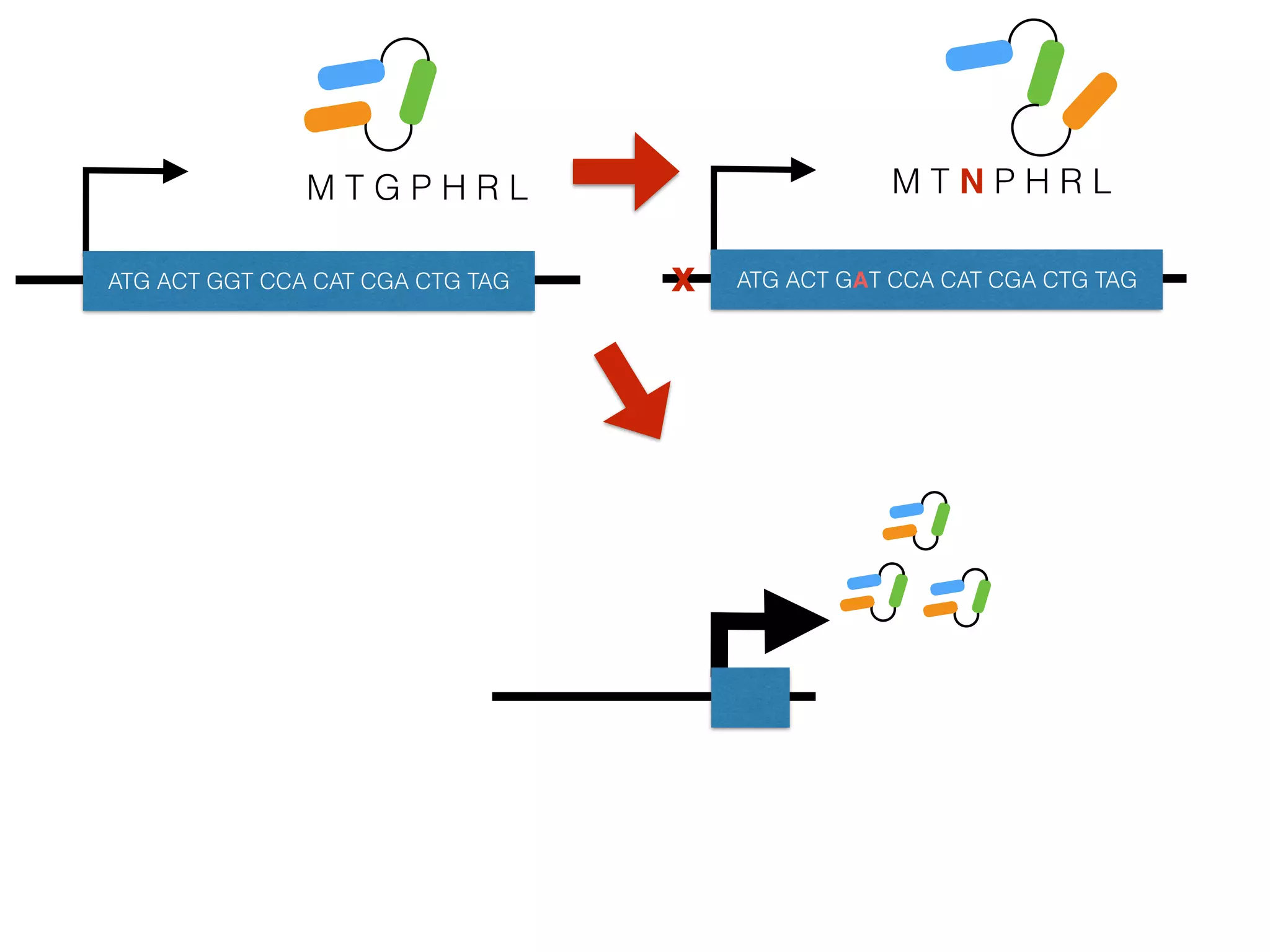
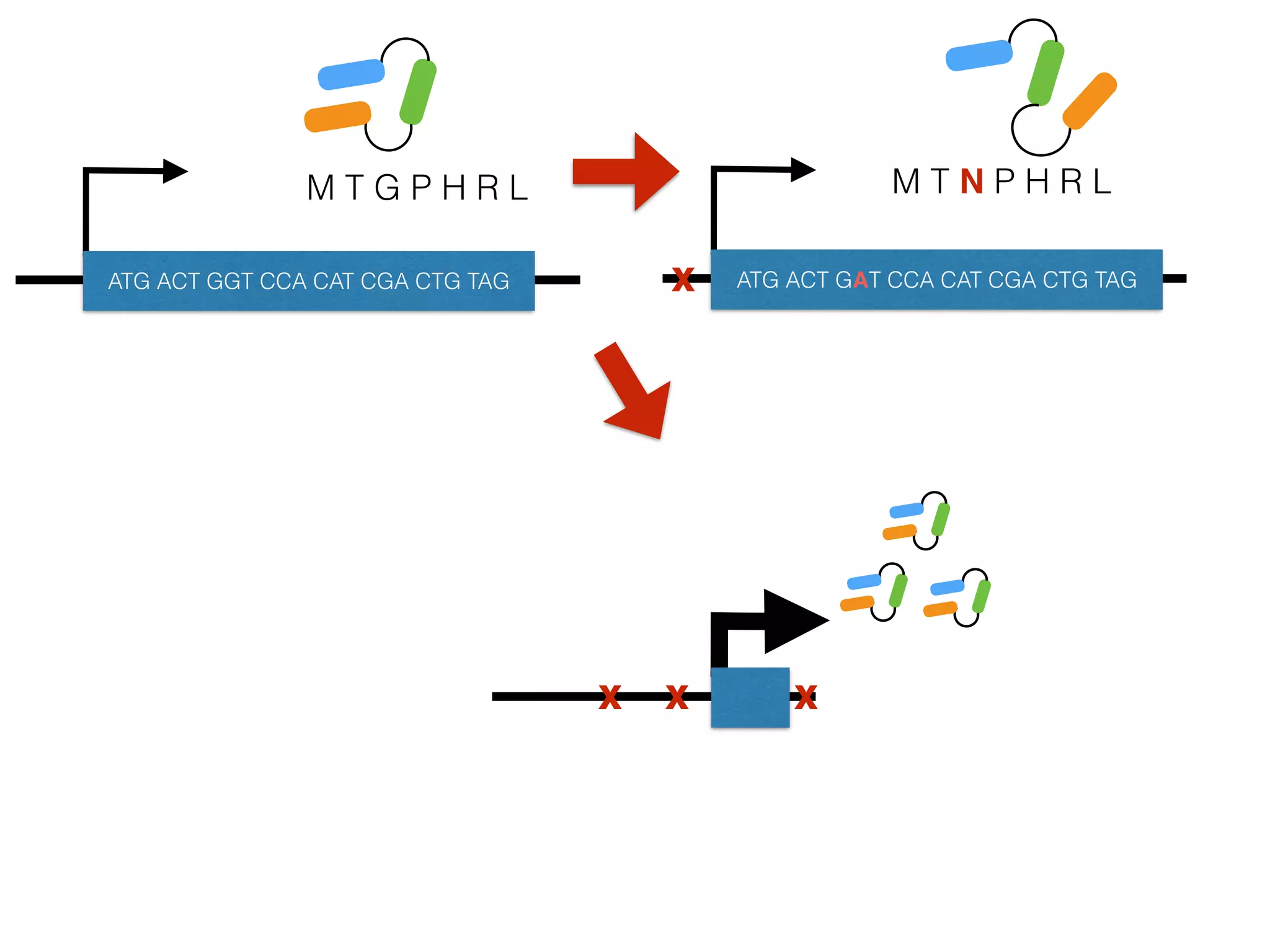
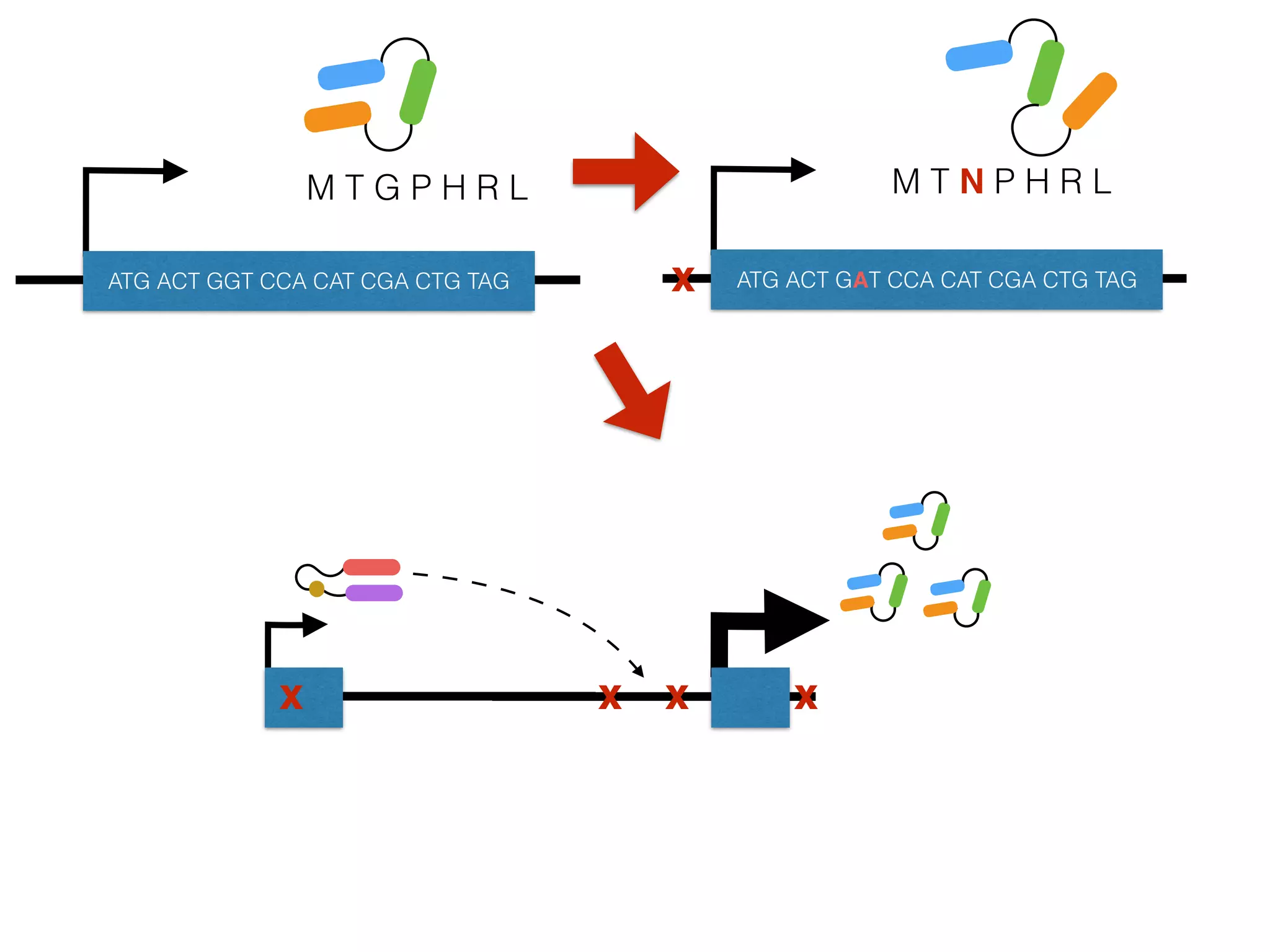
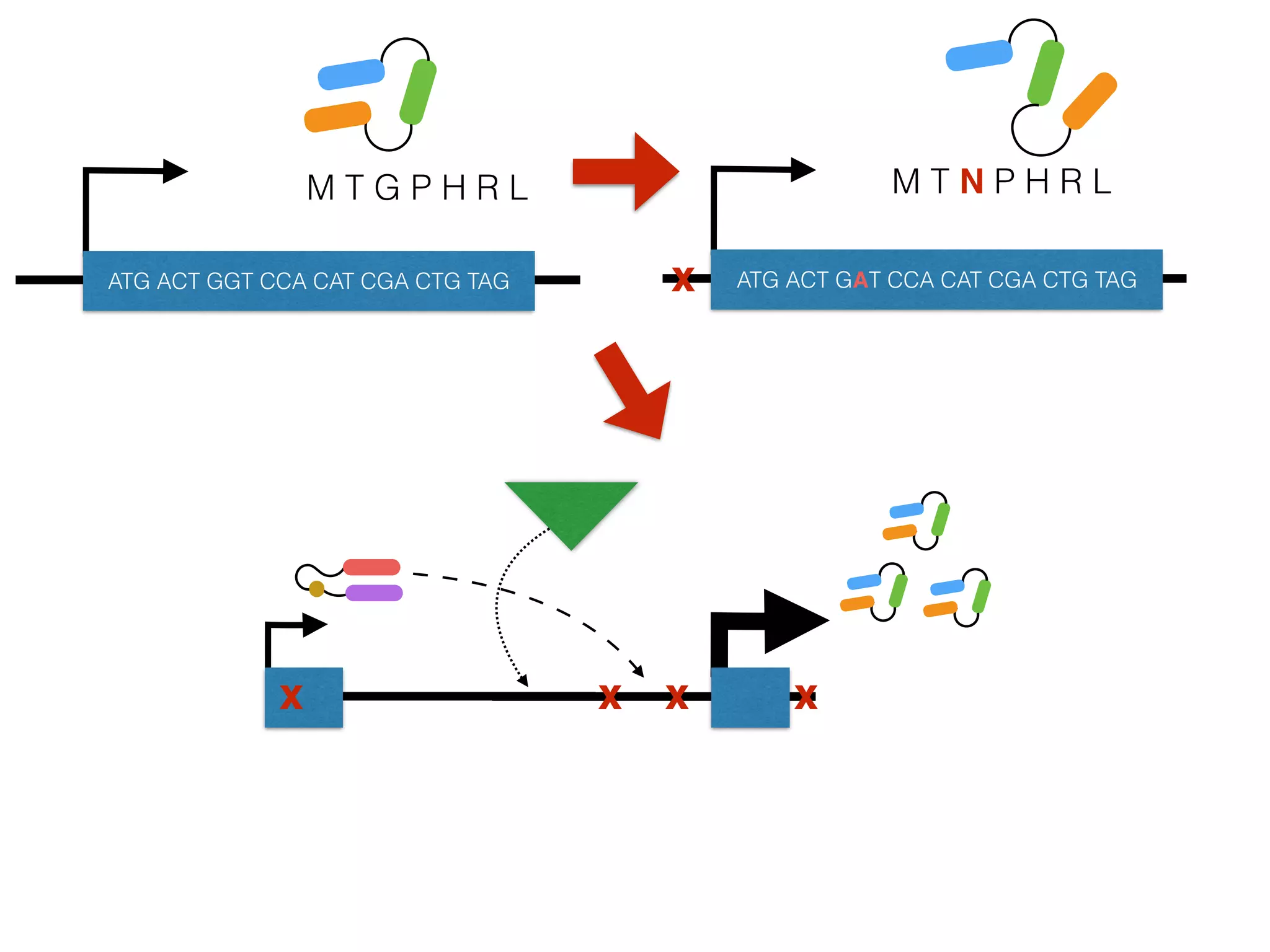

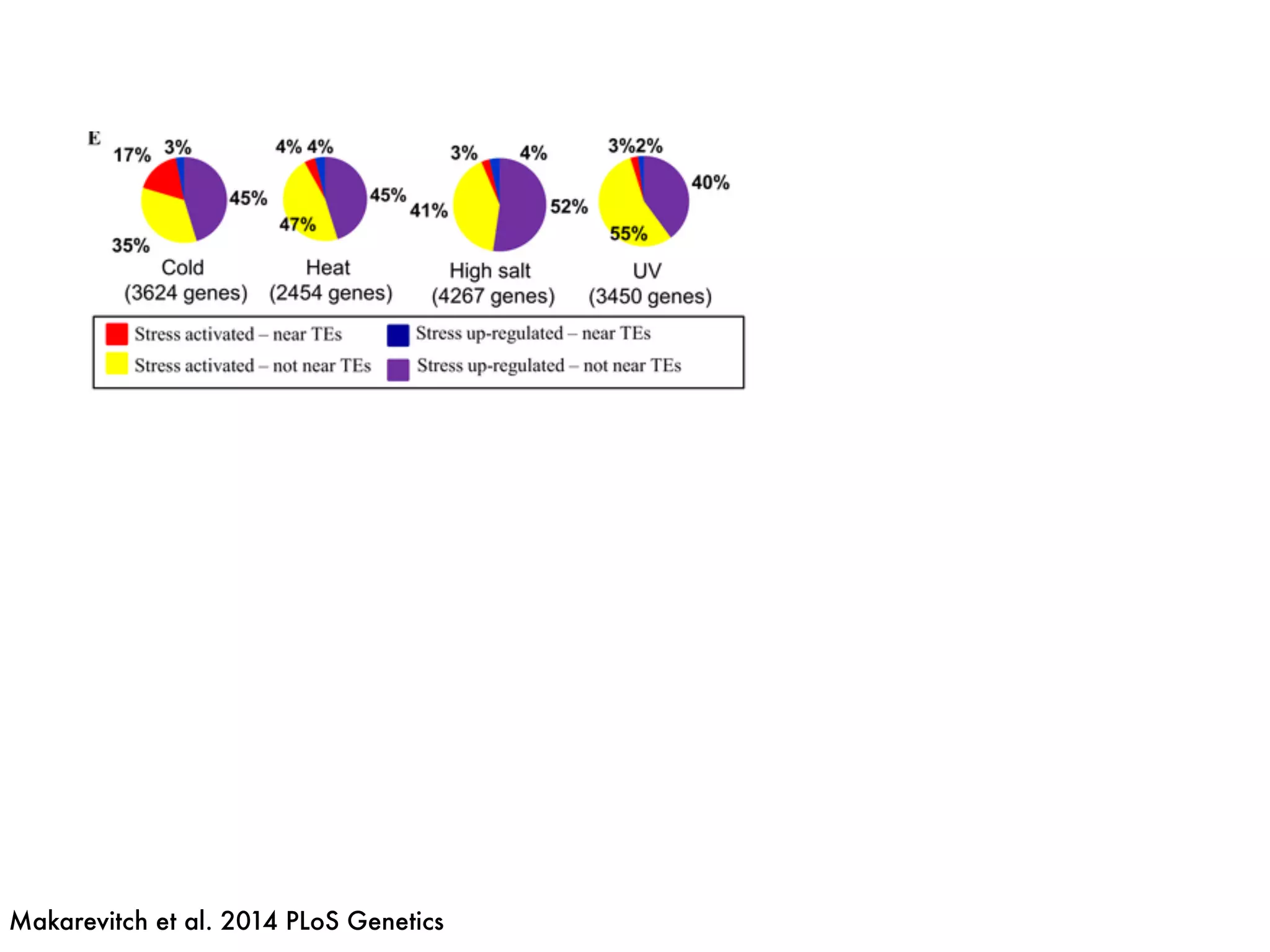
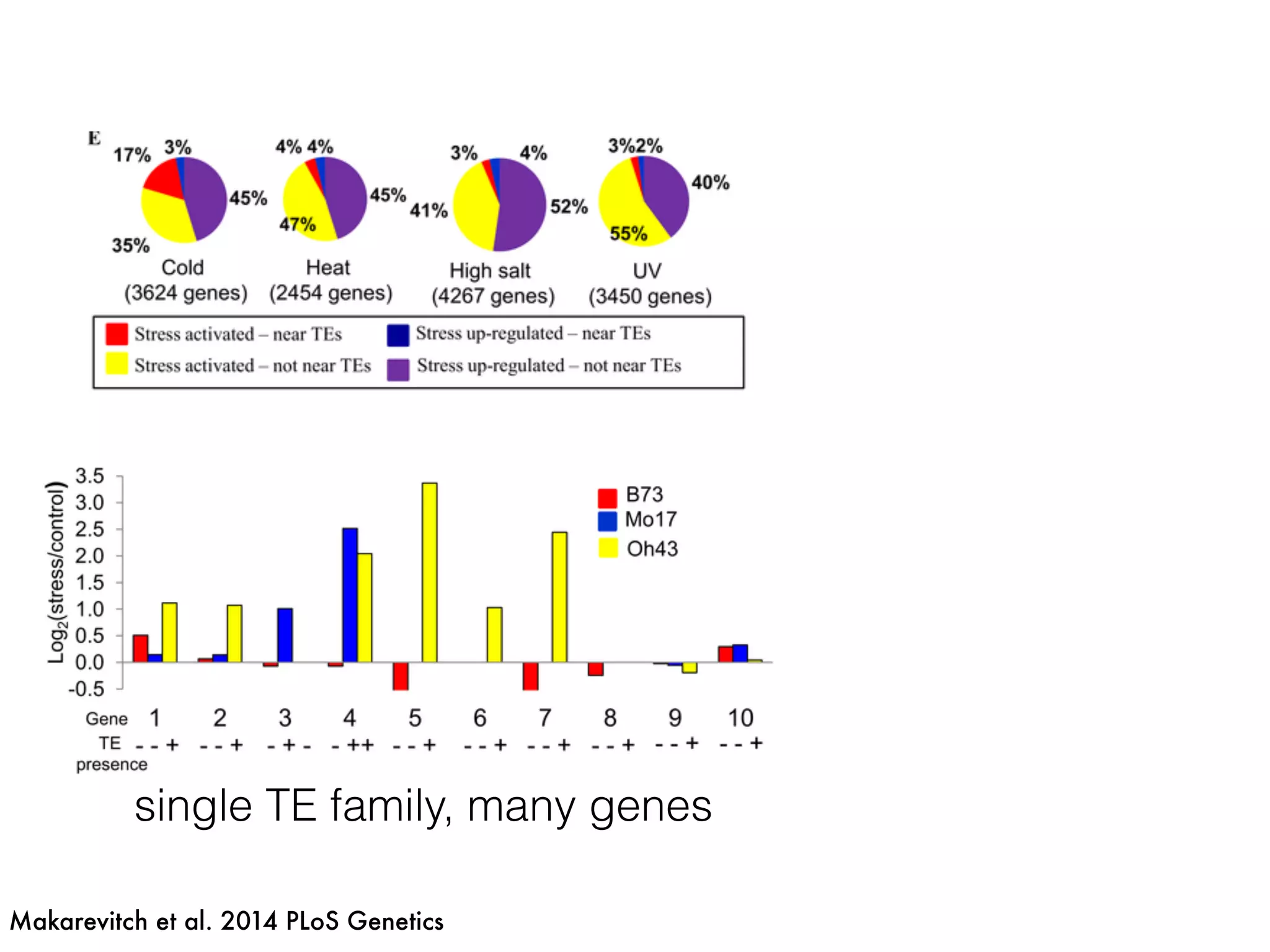
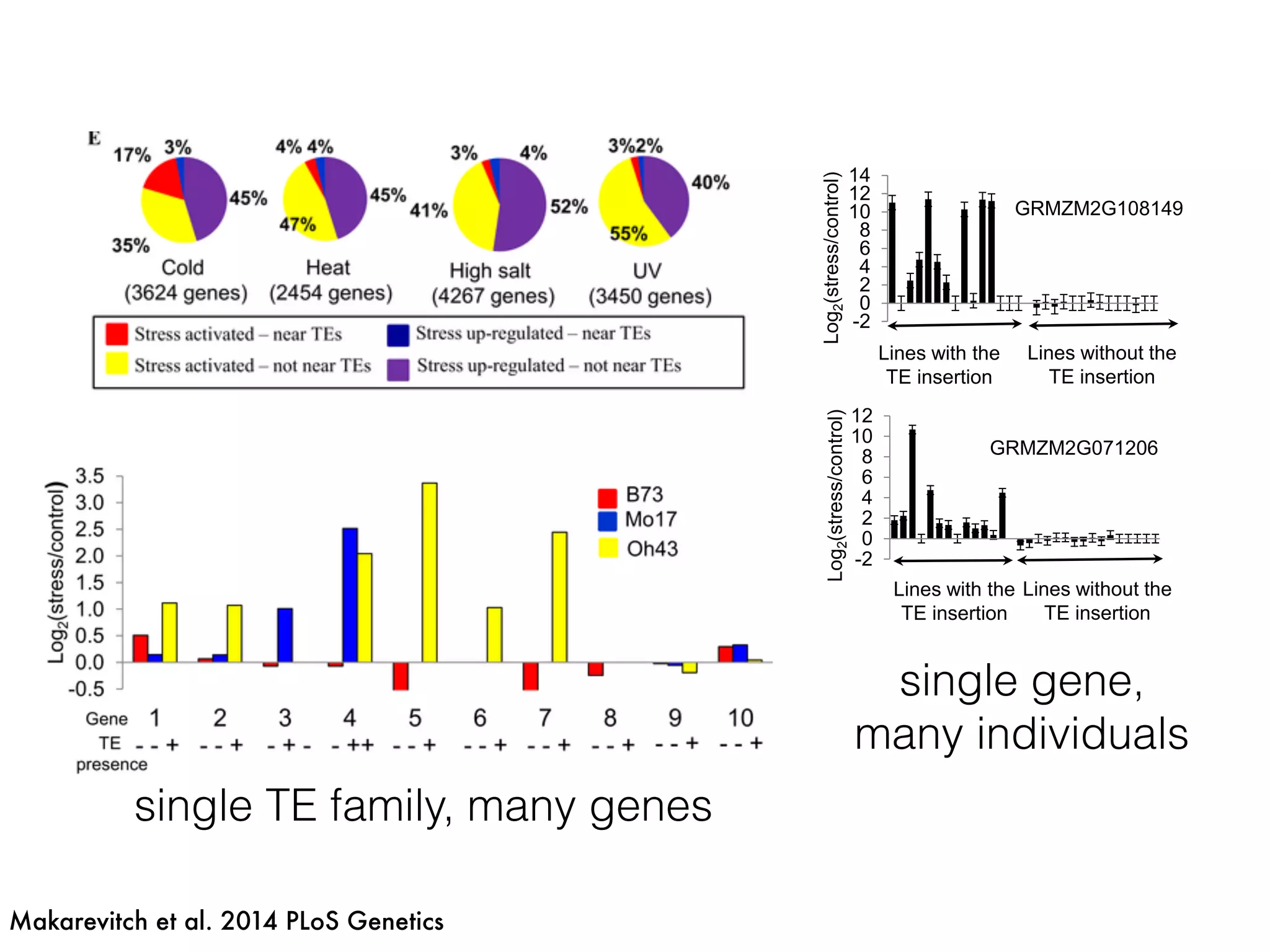
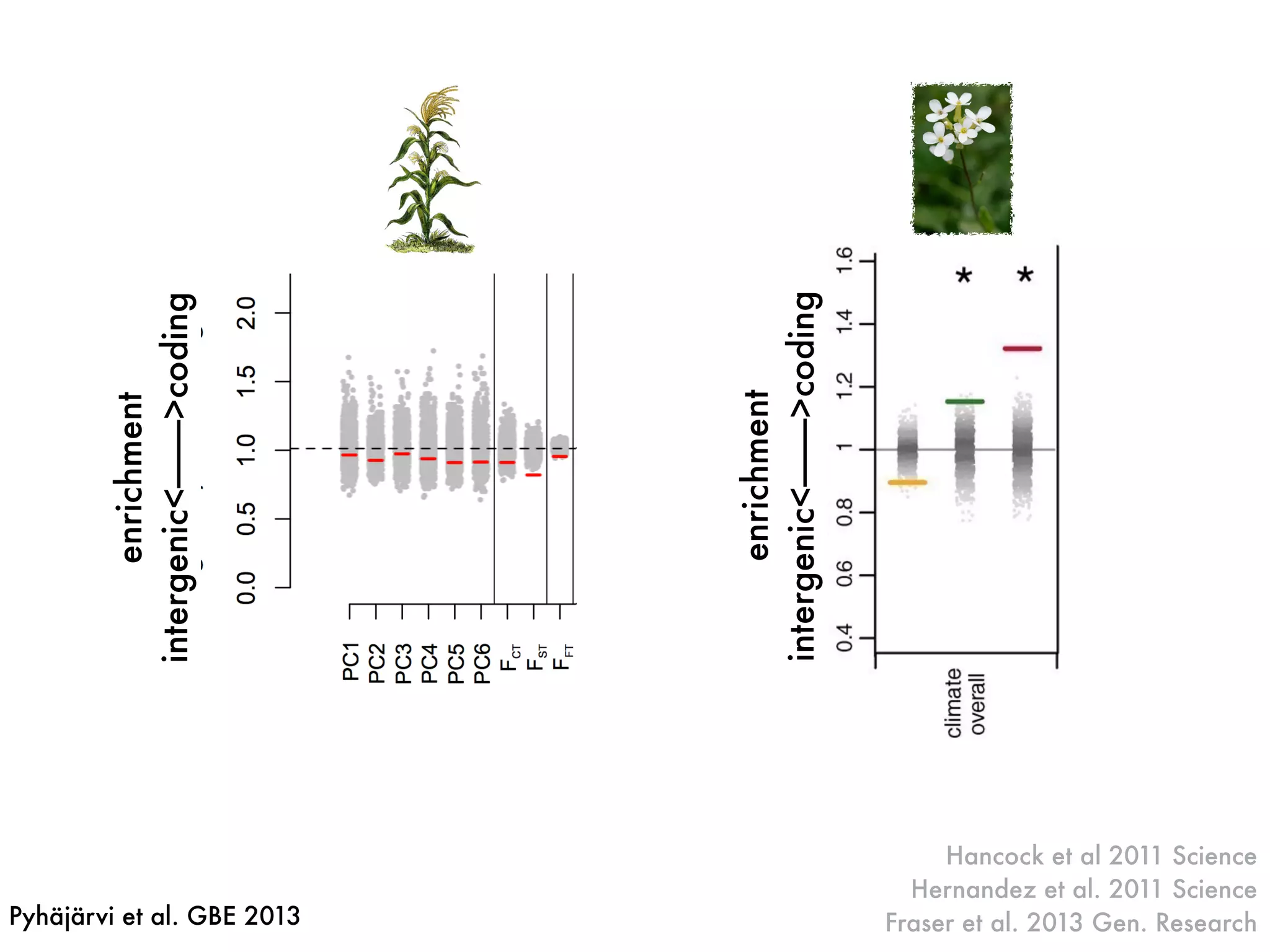



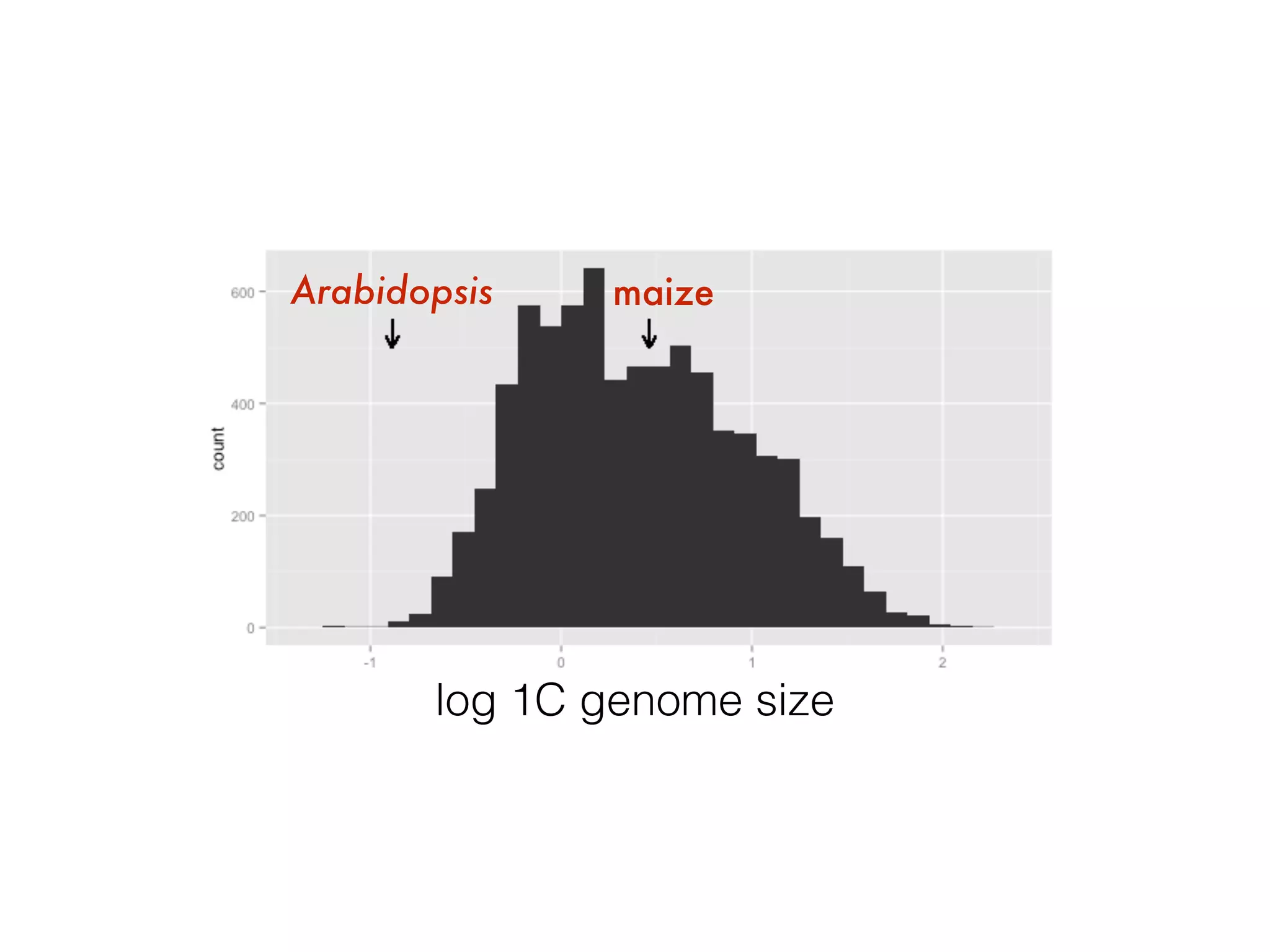































































































The document summarizes research on the selection and demography of maize evolution. It discusses how maize was domesticated from its wild ancestor, teosinte, through selection on specific genes like tga1 and tb1 that contributed to changes in plant architecture. It also describes the spread of maize from its center of domestication in Mexico to other regions, noting evidence of selection during this spread. Genomic analyses reveal thousands of genes underwent changes between teosinte and modern maize varieties through both hard sweeps on individual genes and polygenic adaptation.


























































































